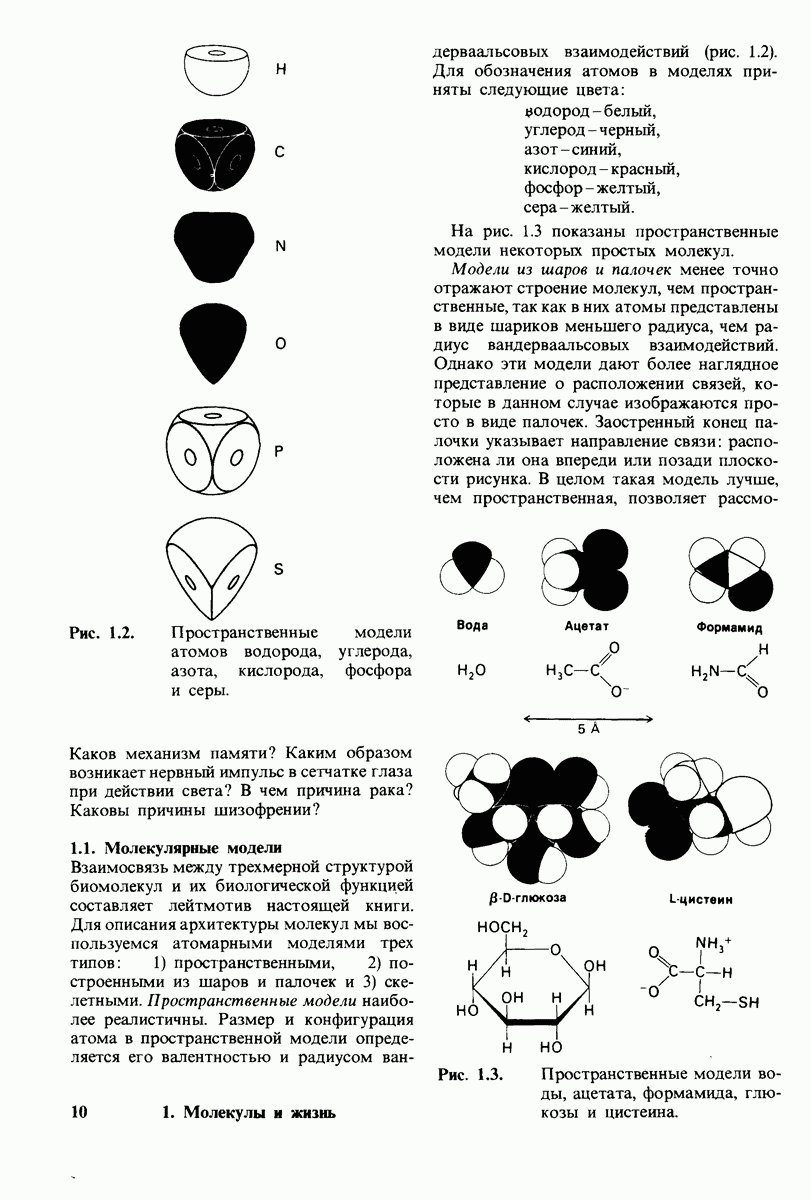
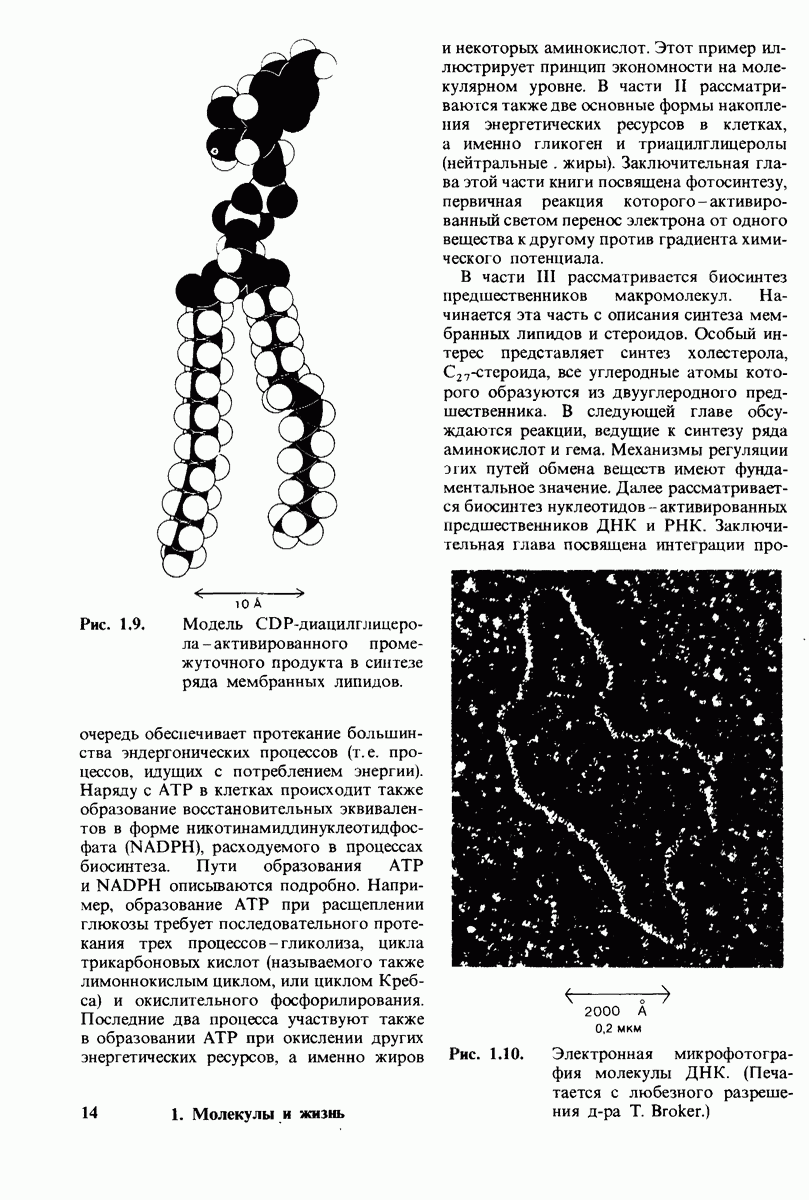
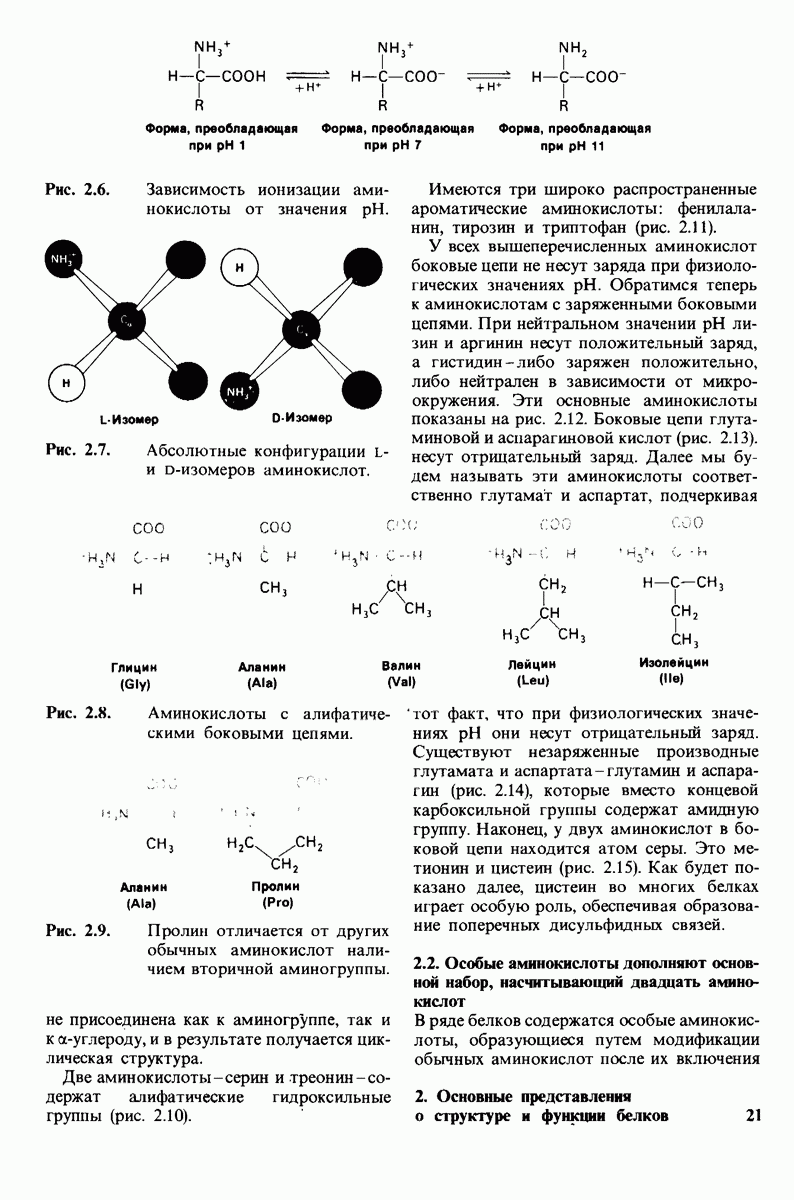
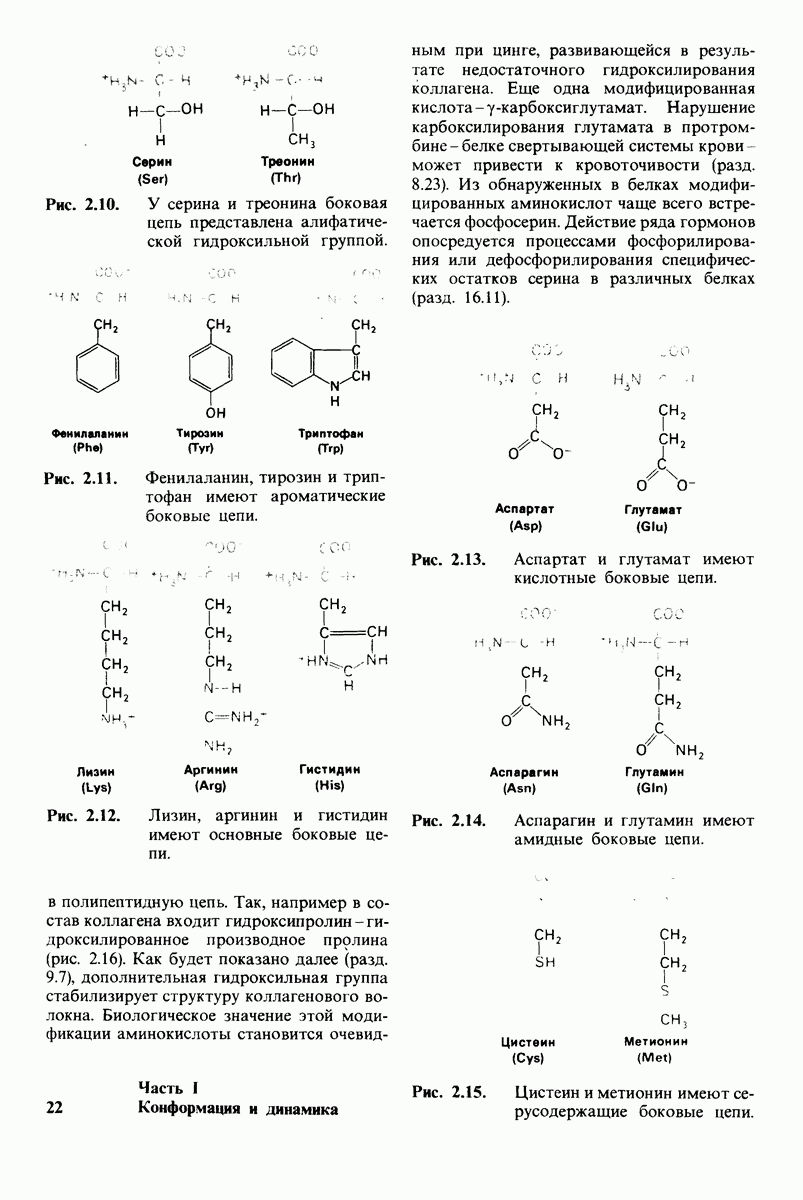
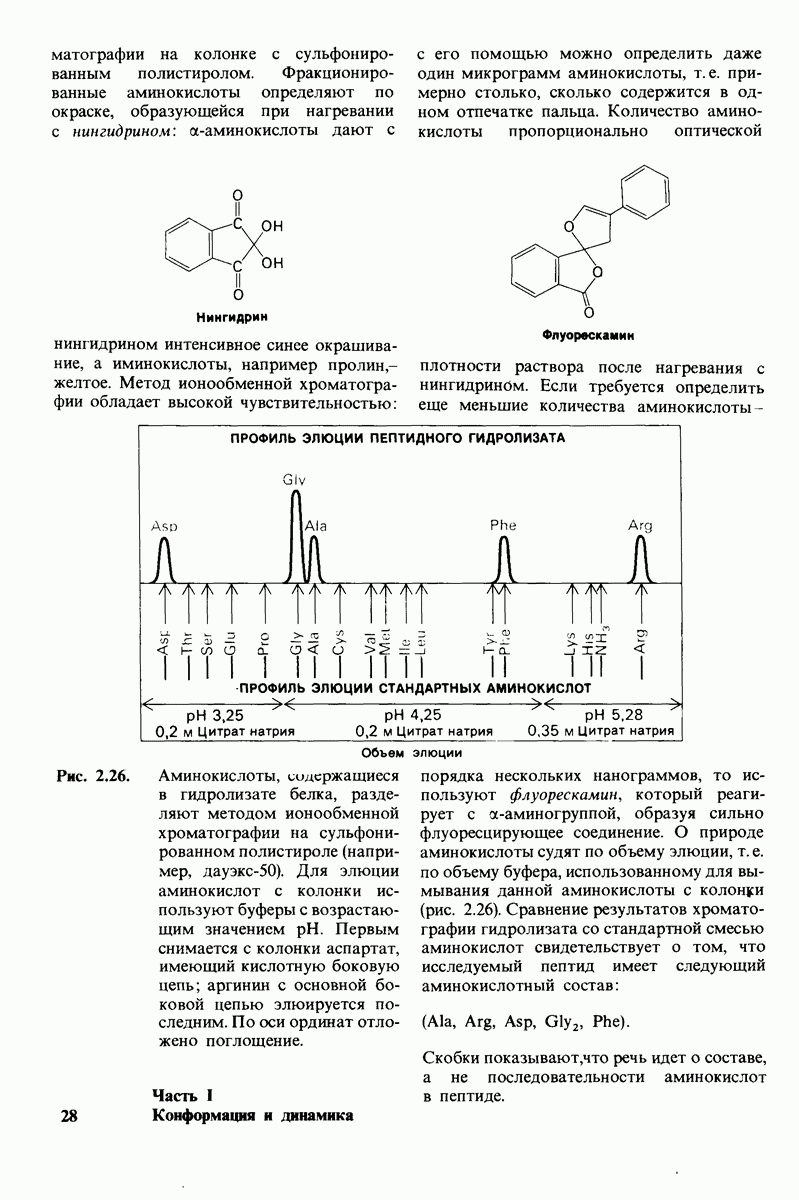
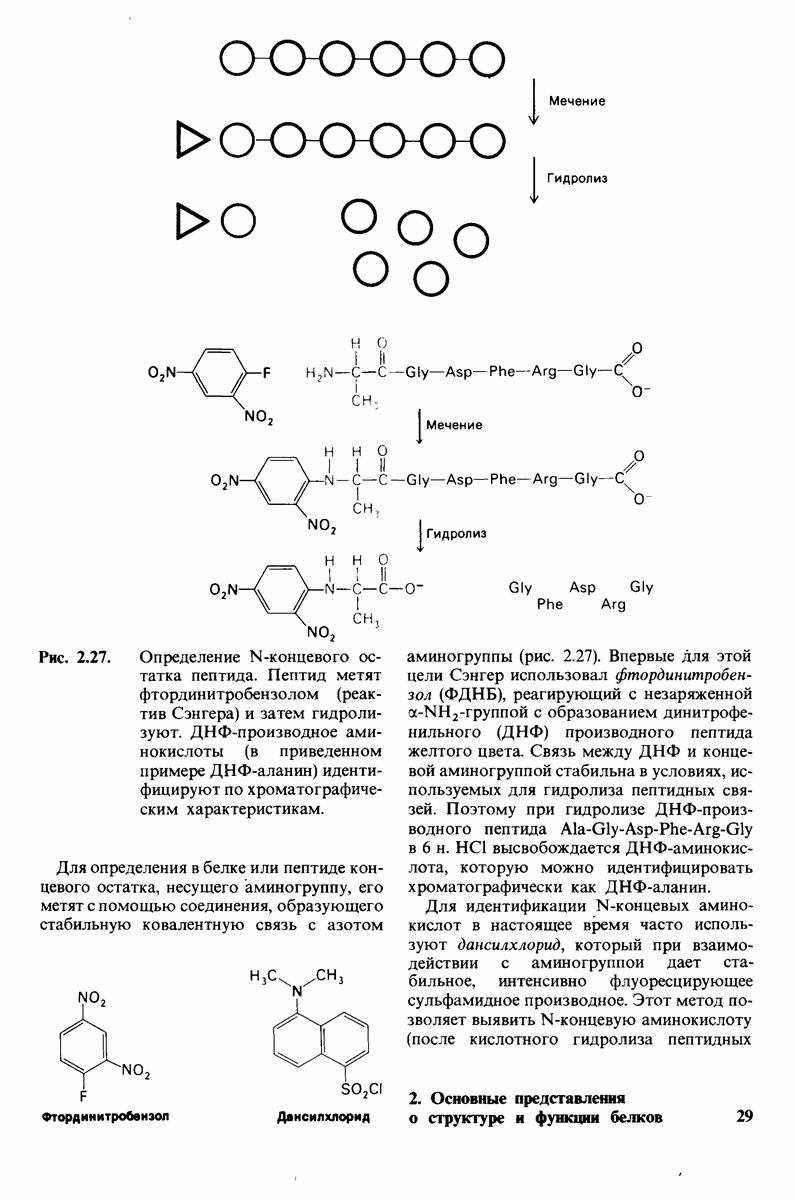
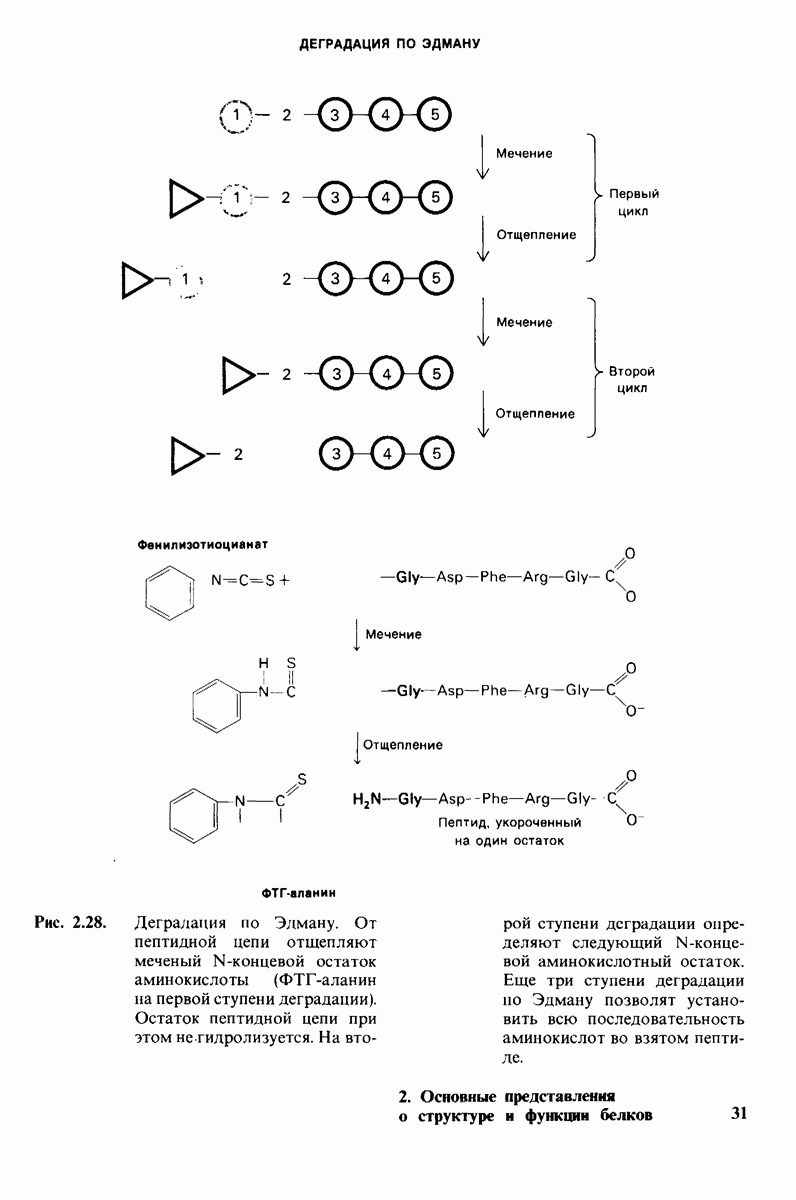
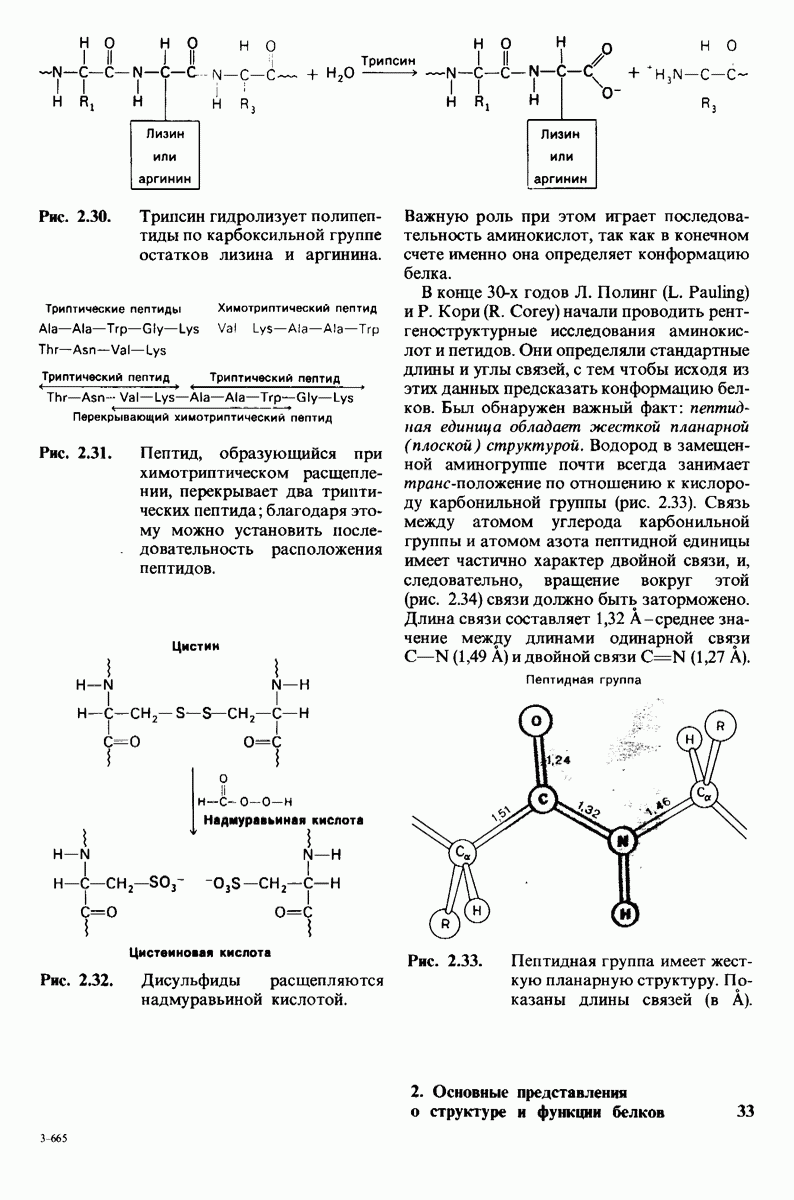
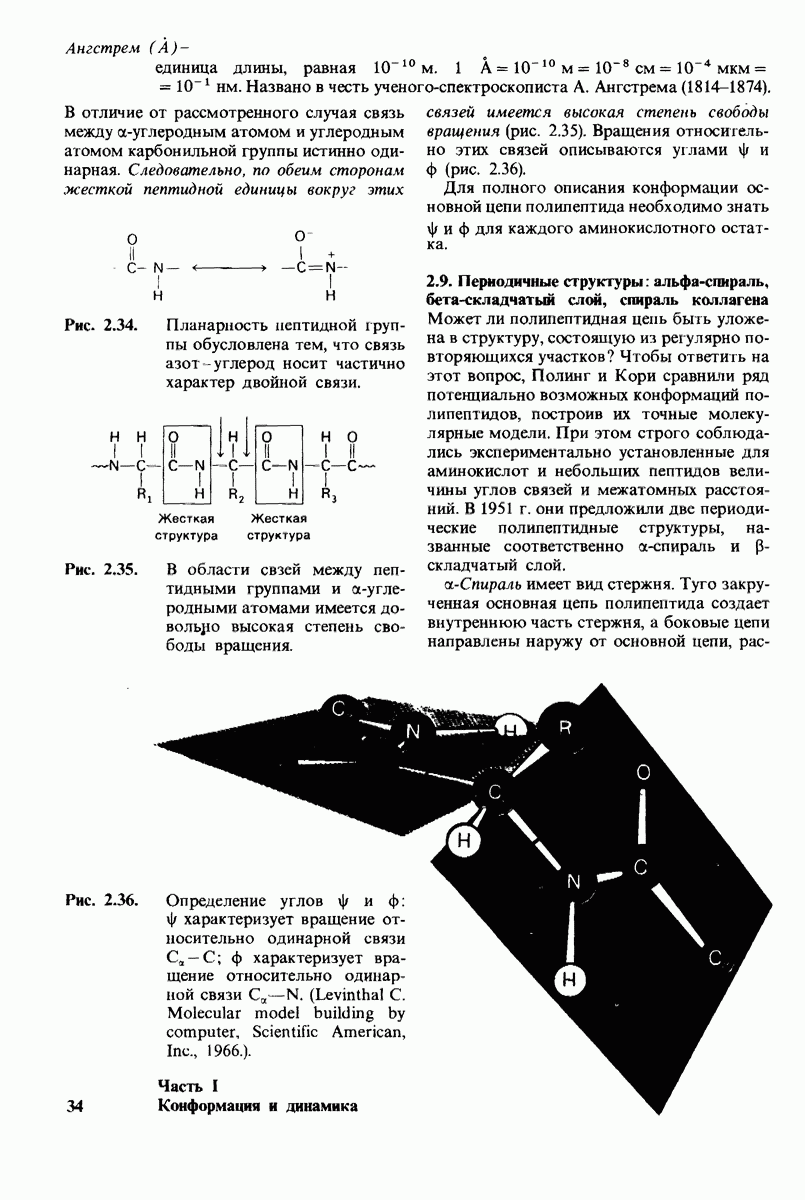
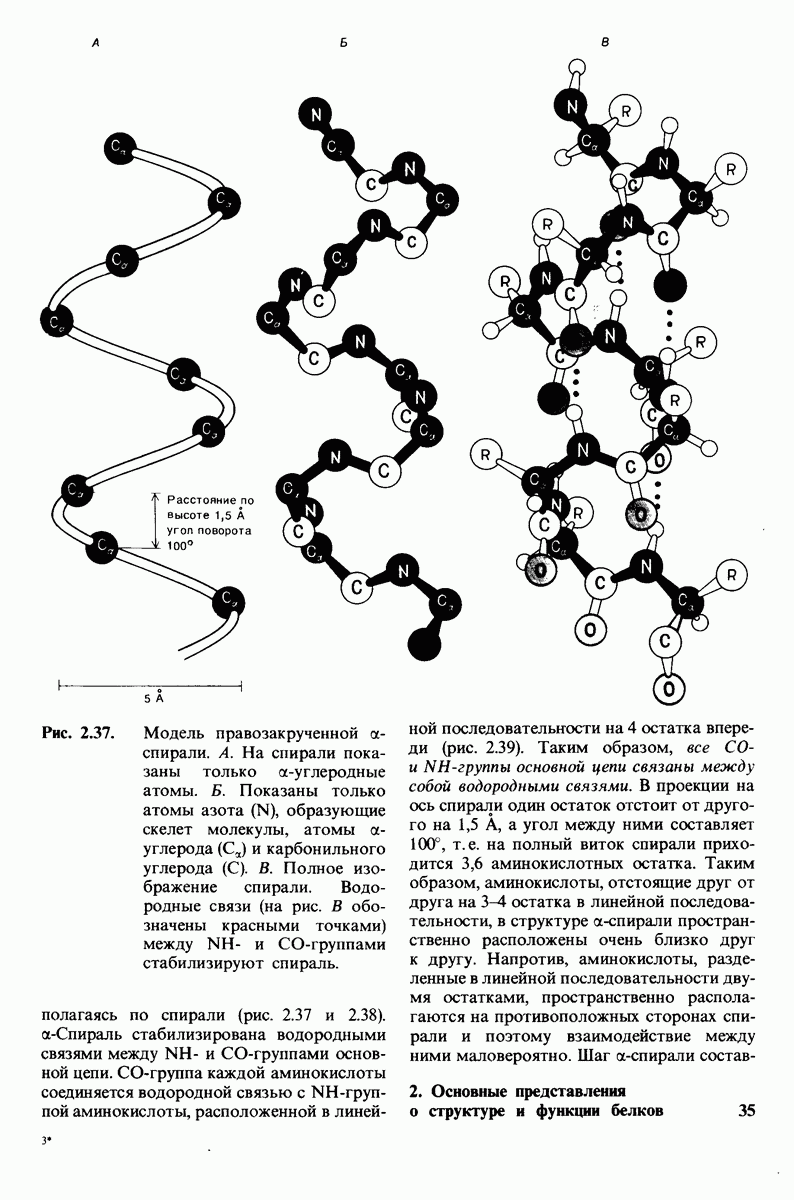
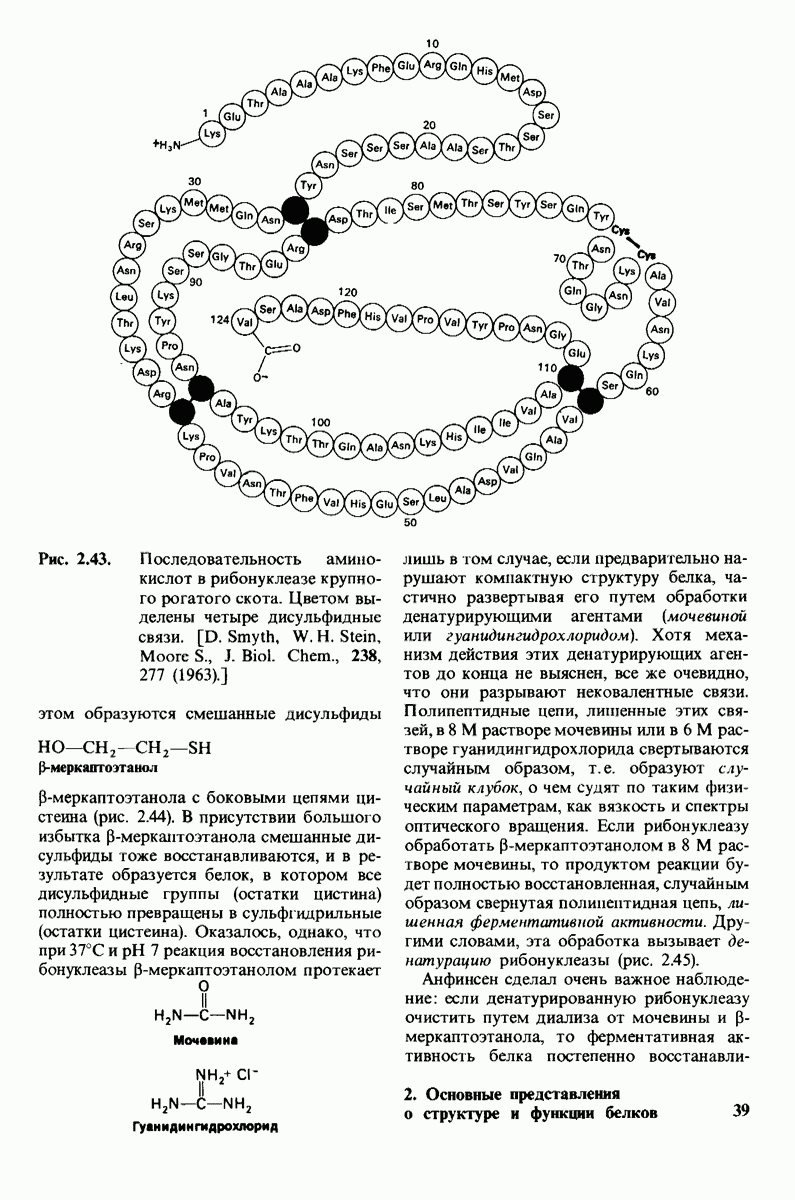
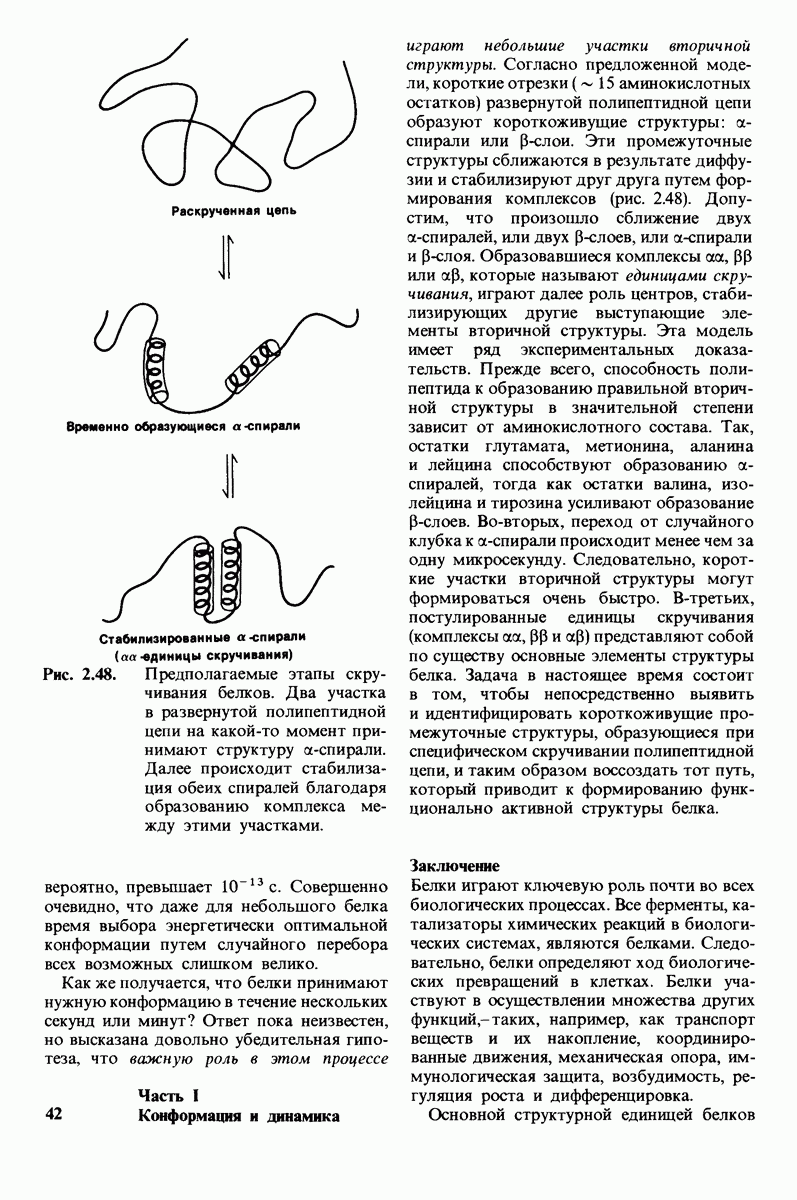
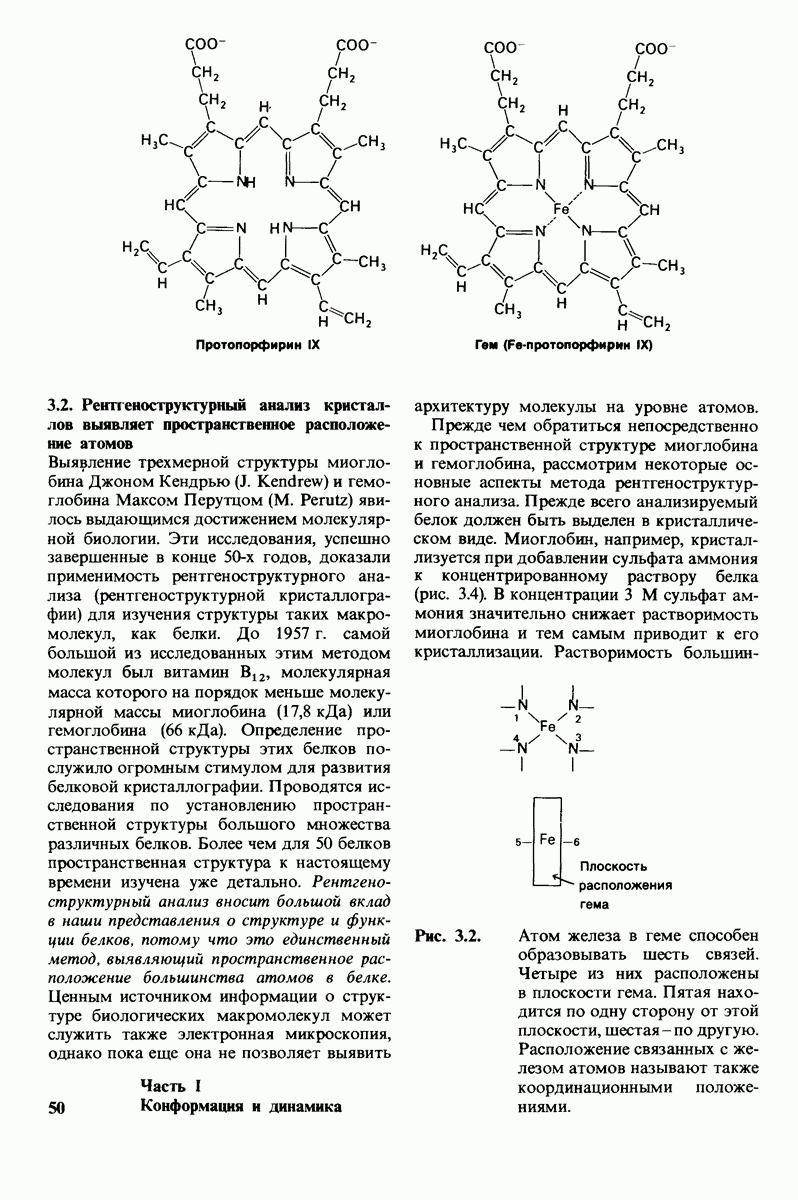
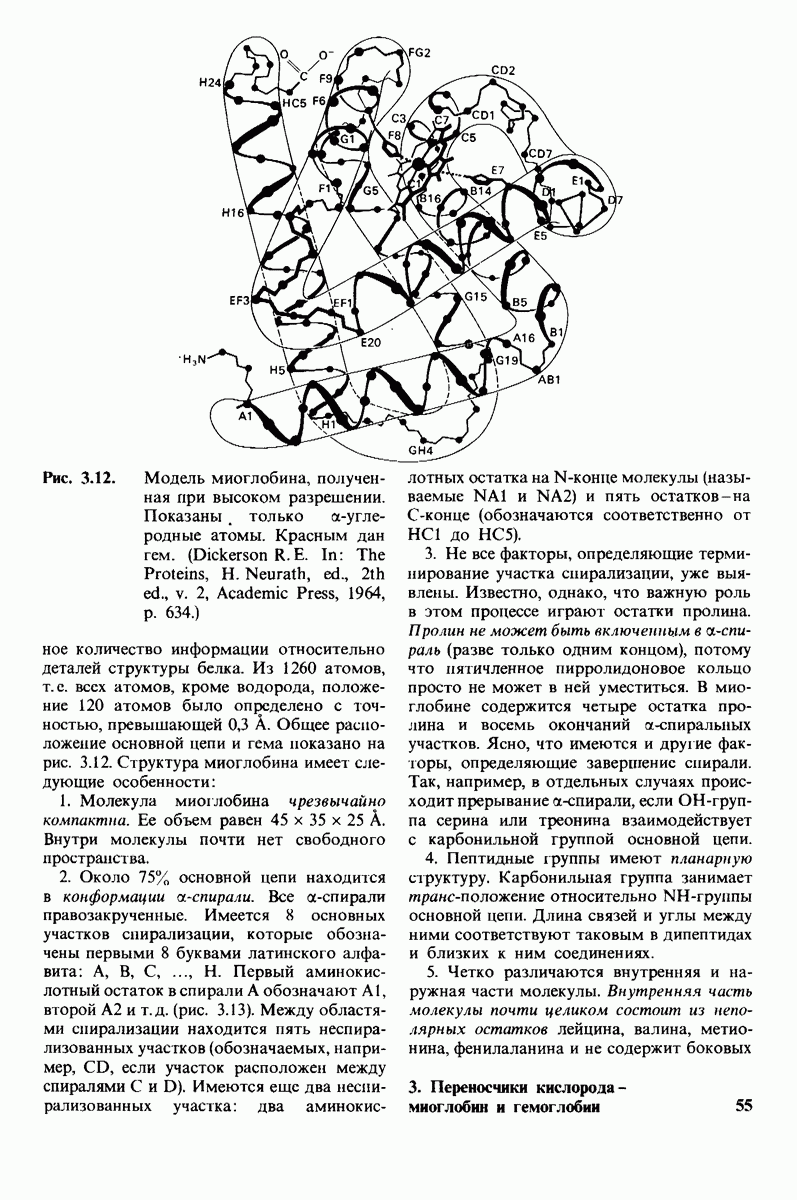
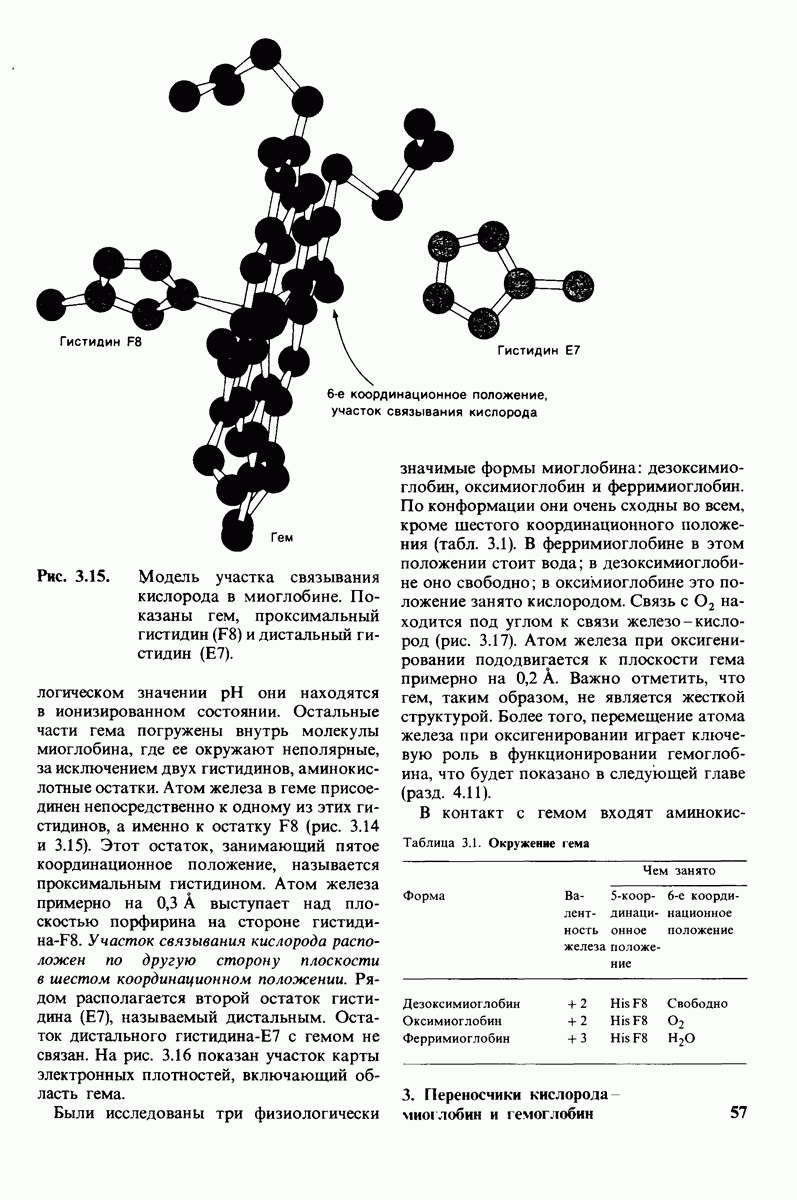
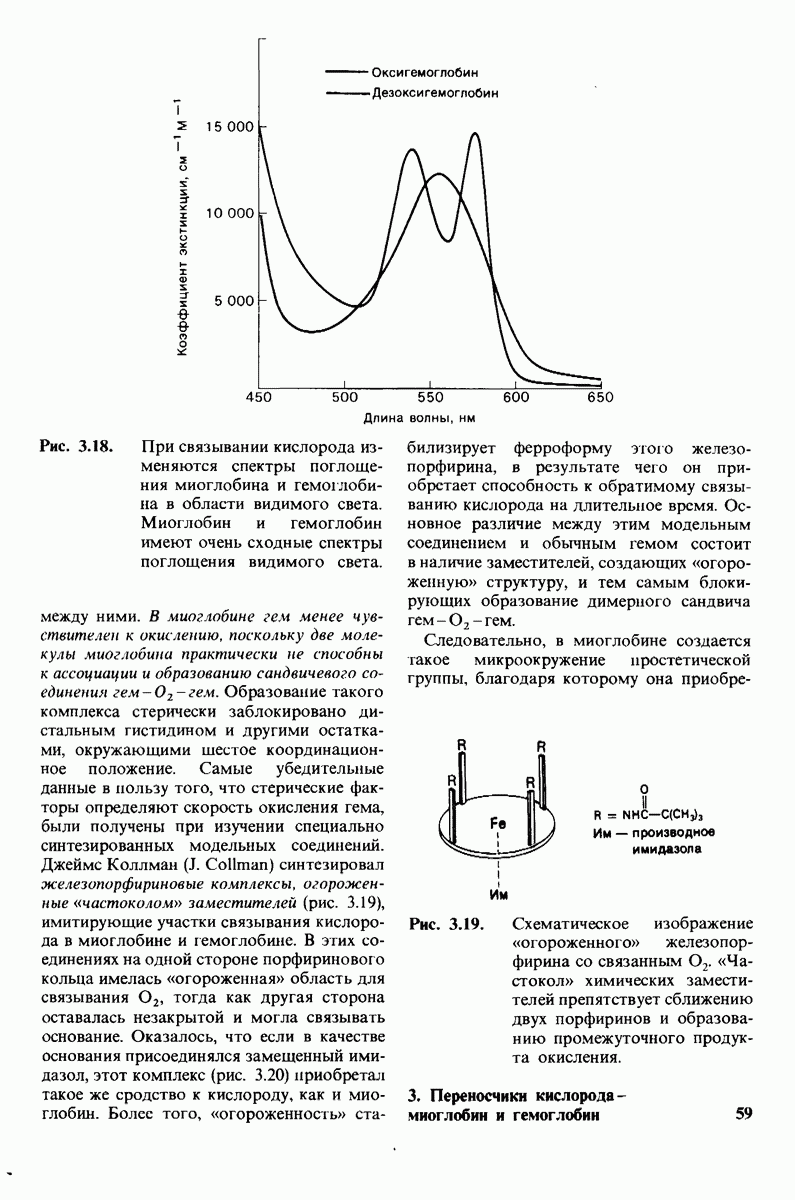
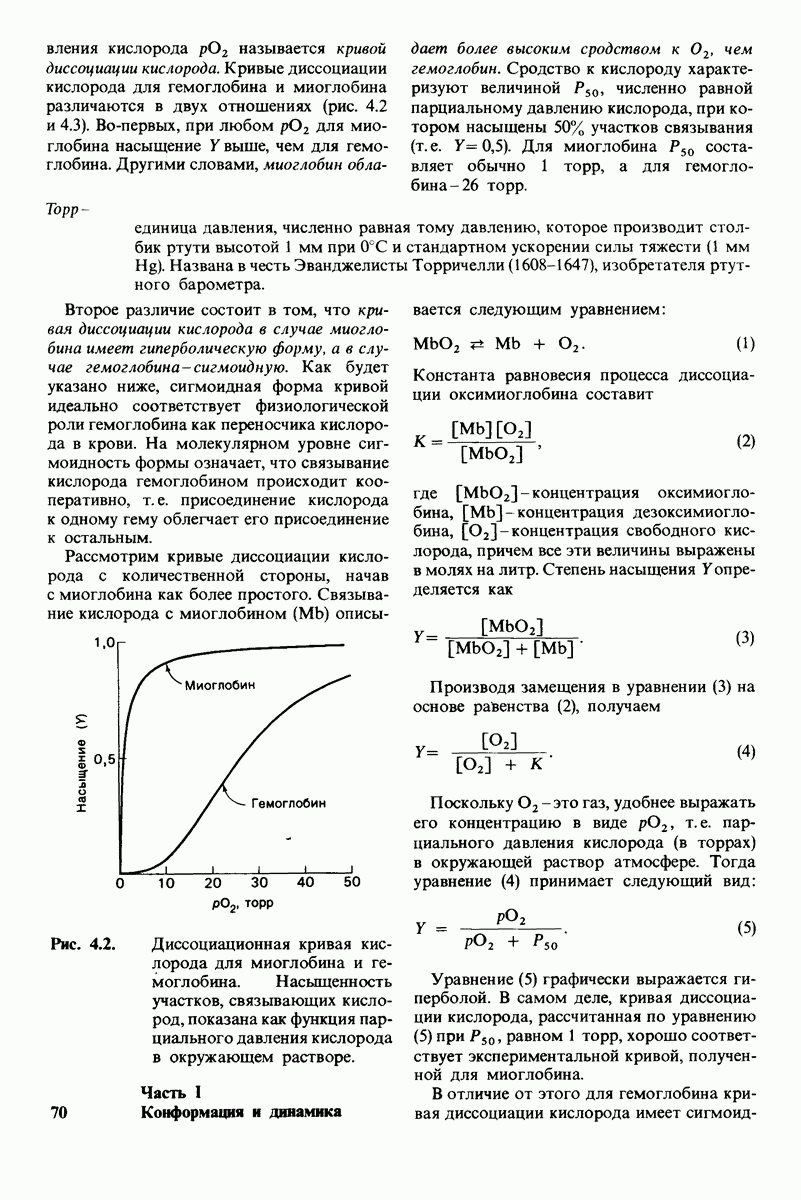
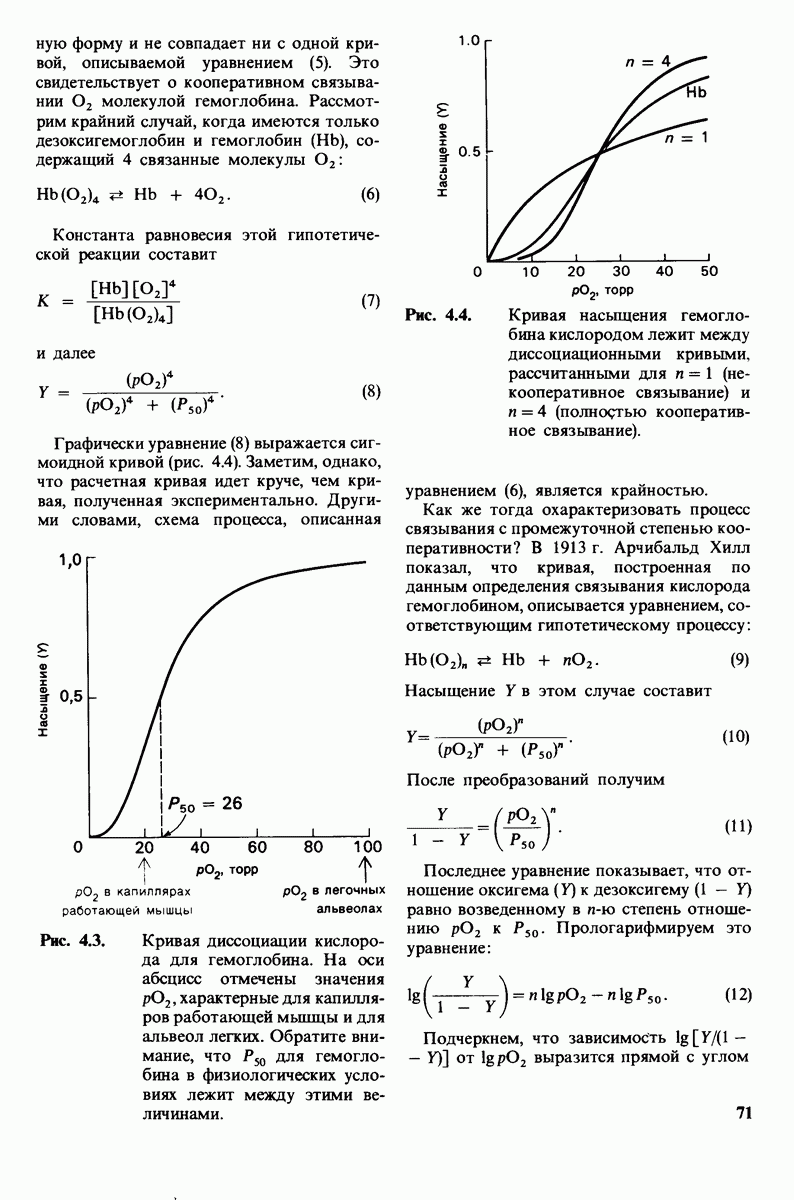
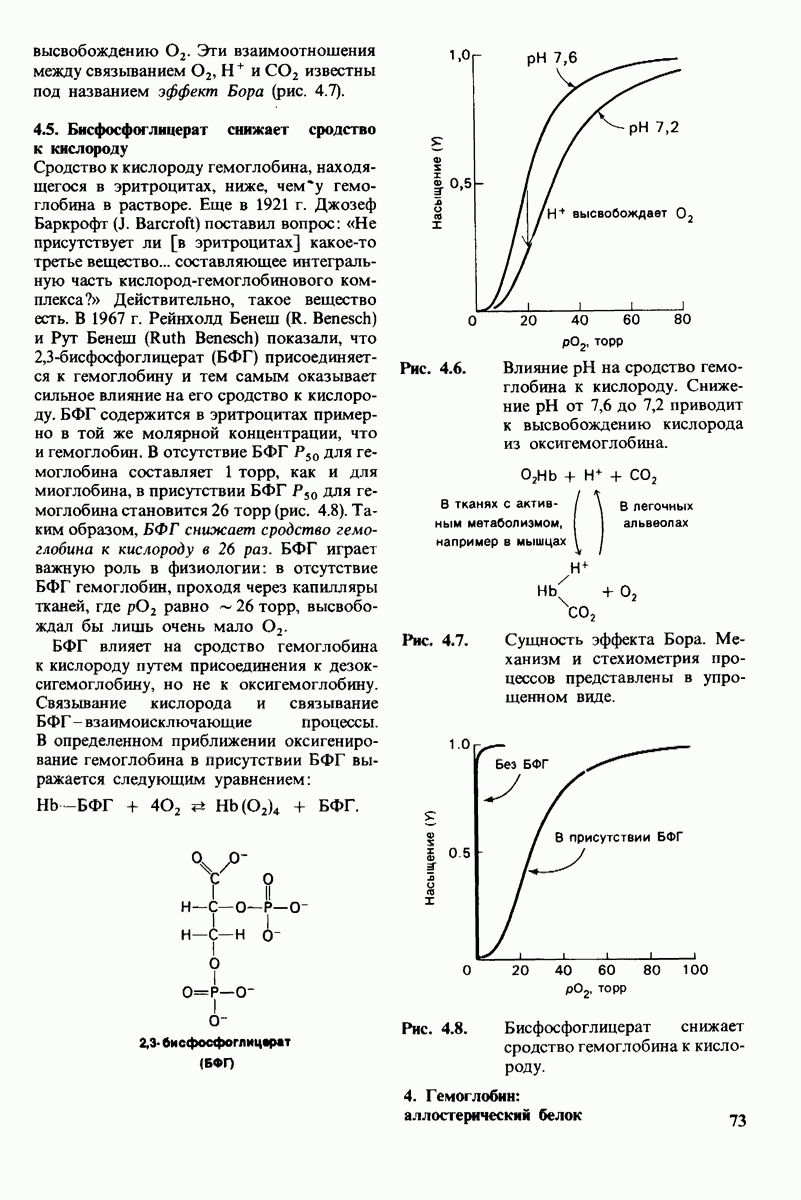
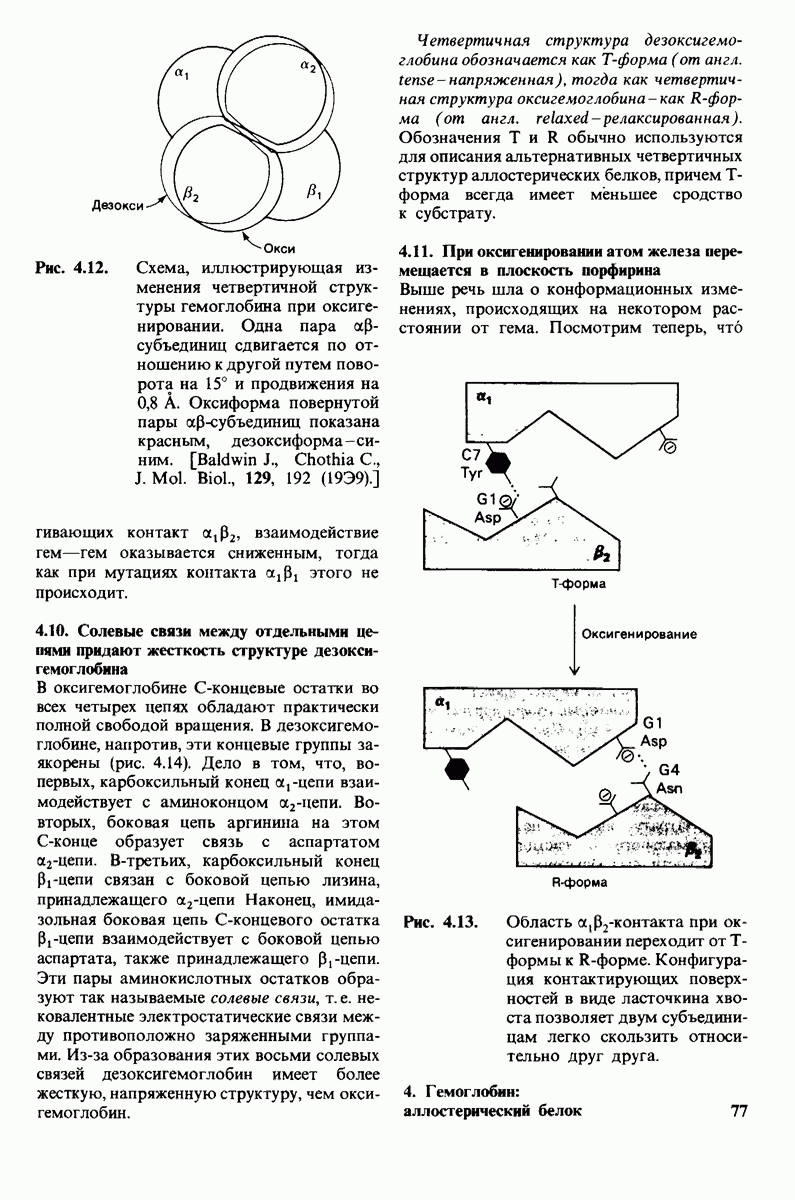
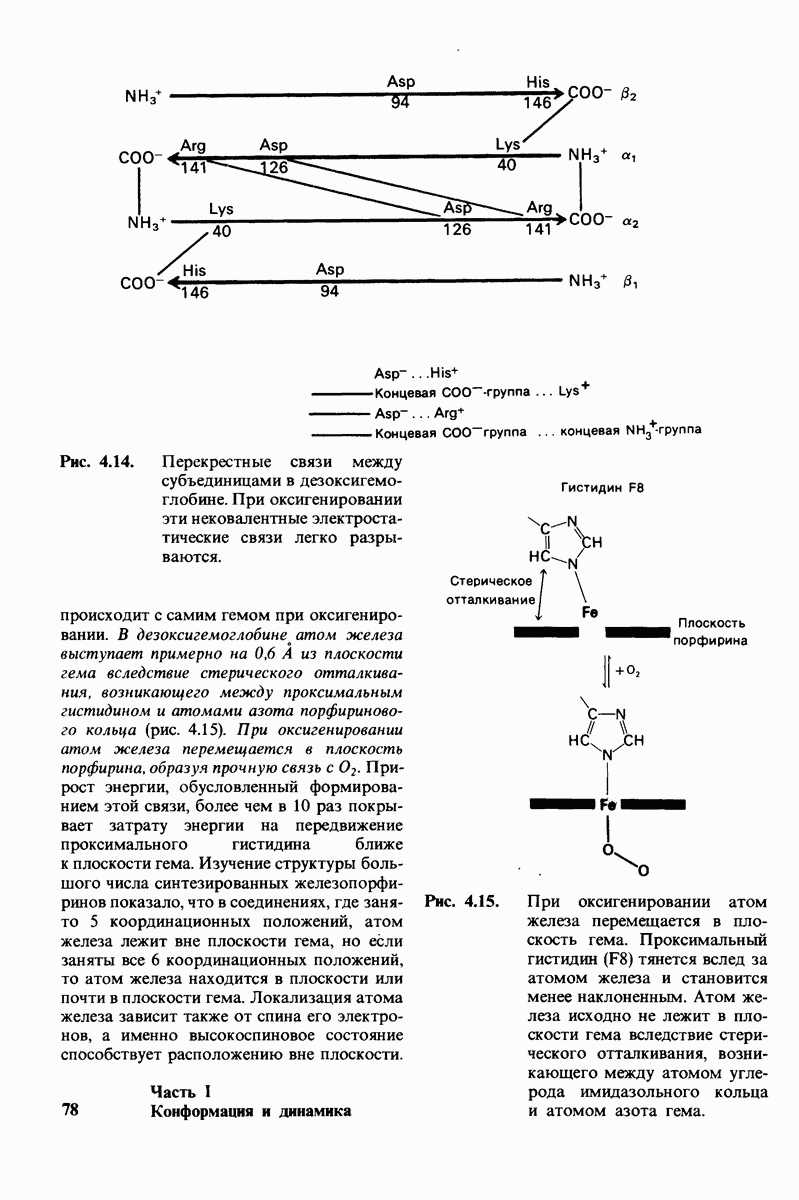
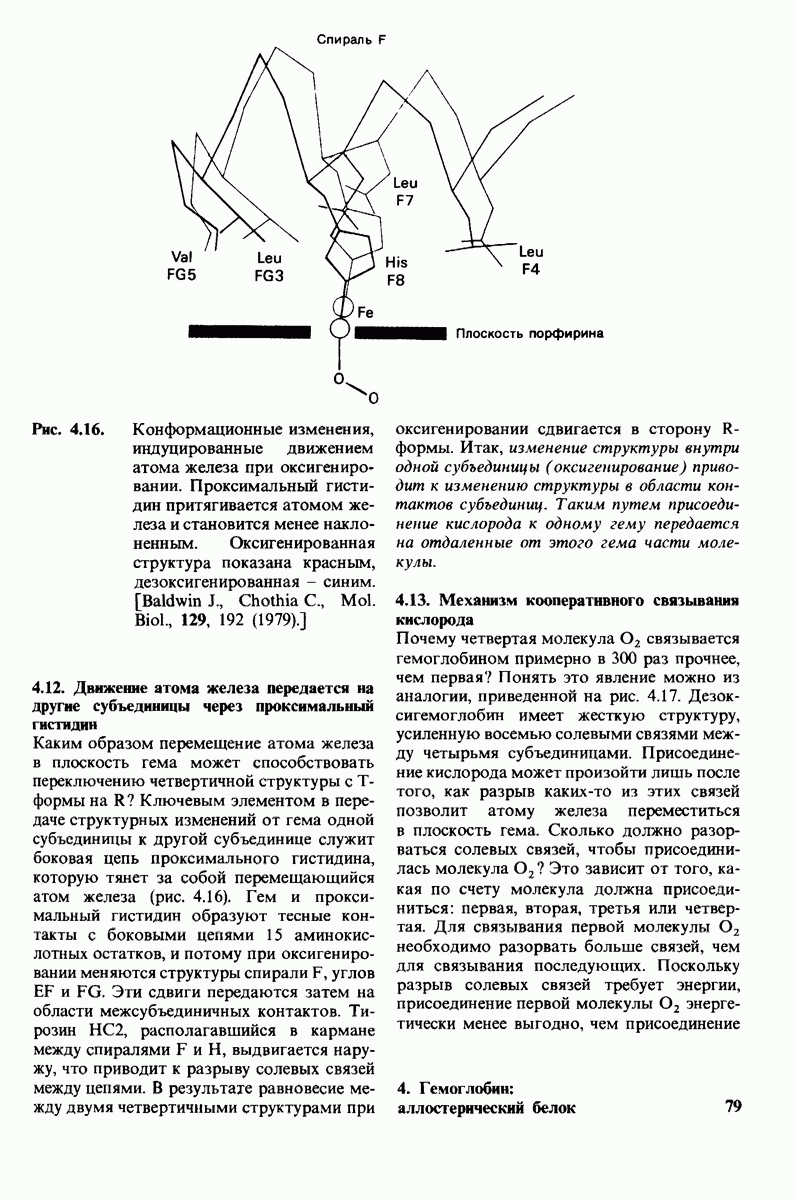
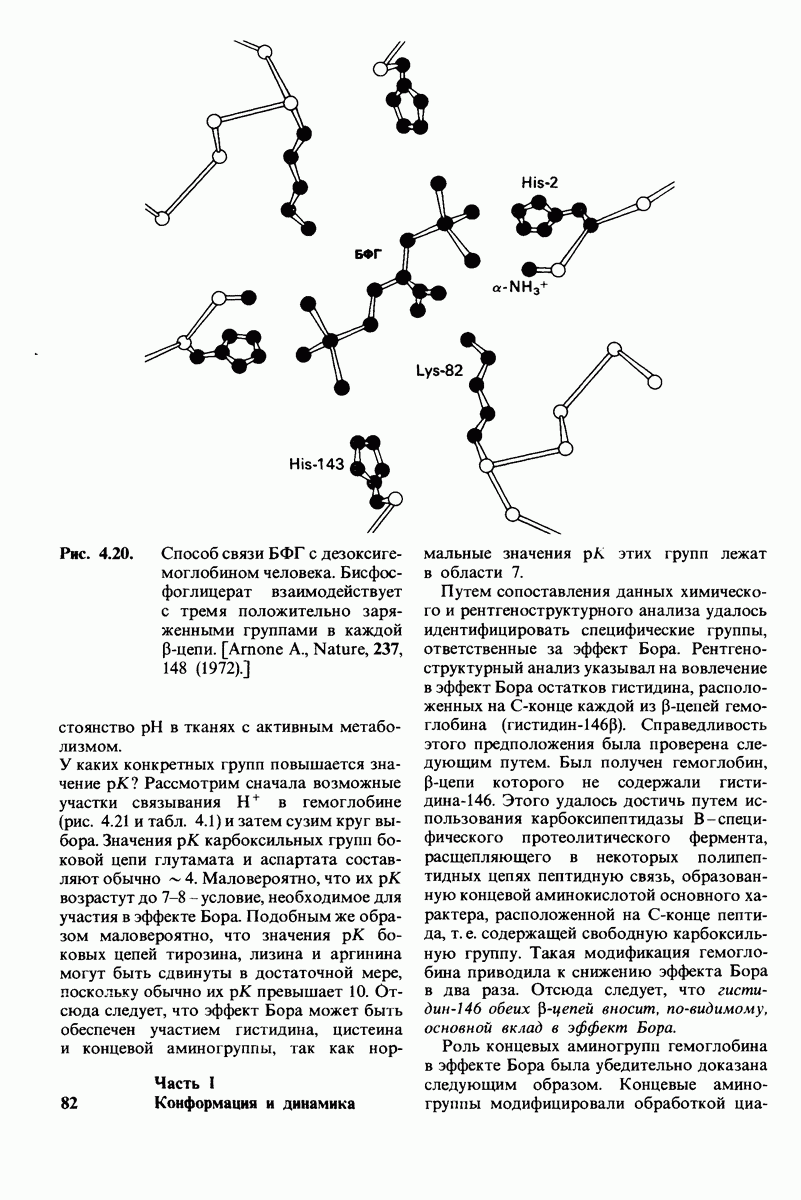
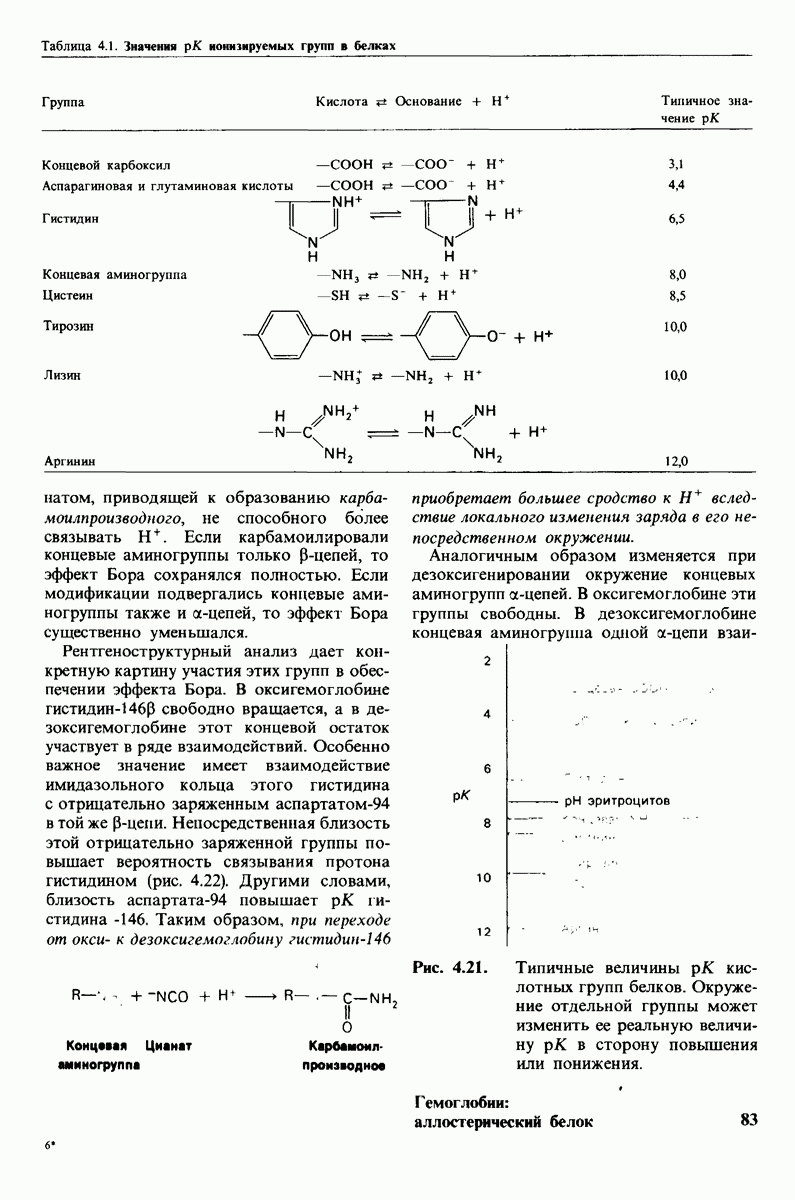
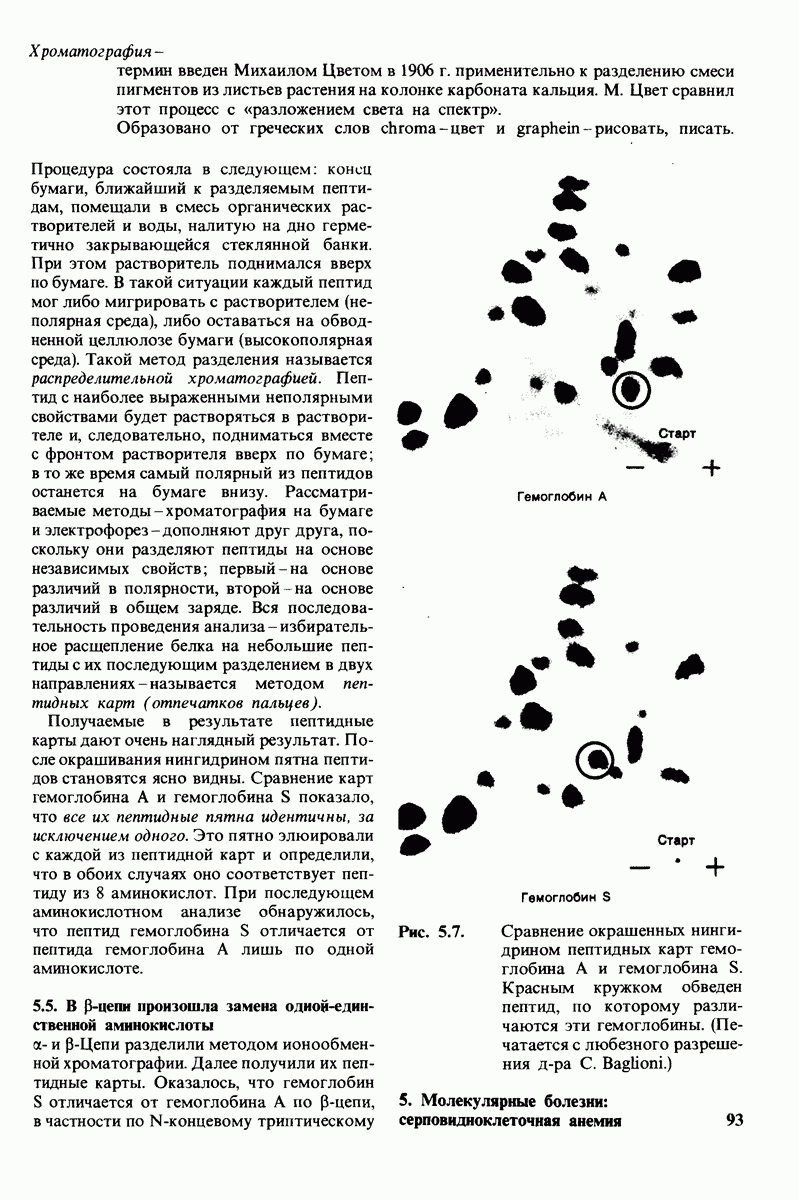
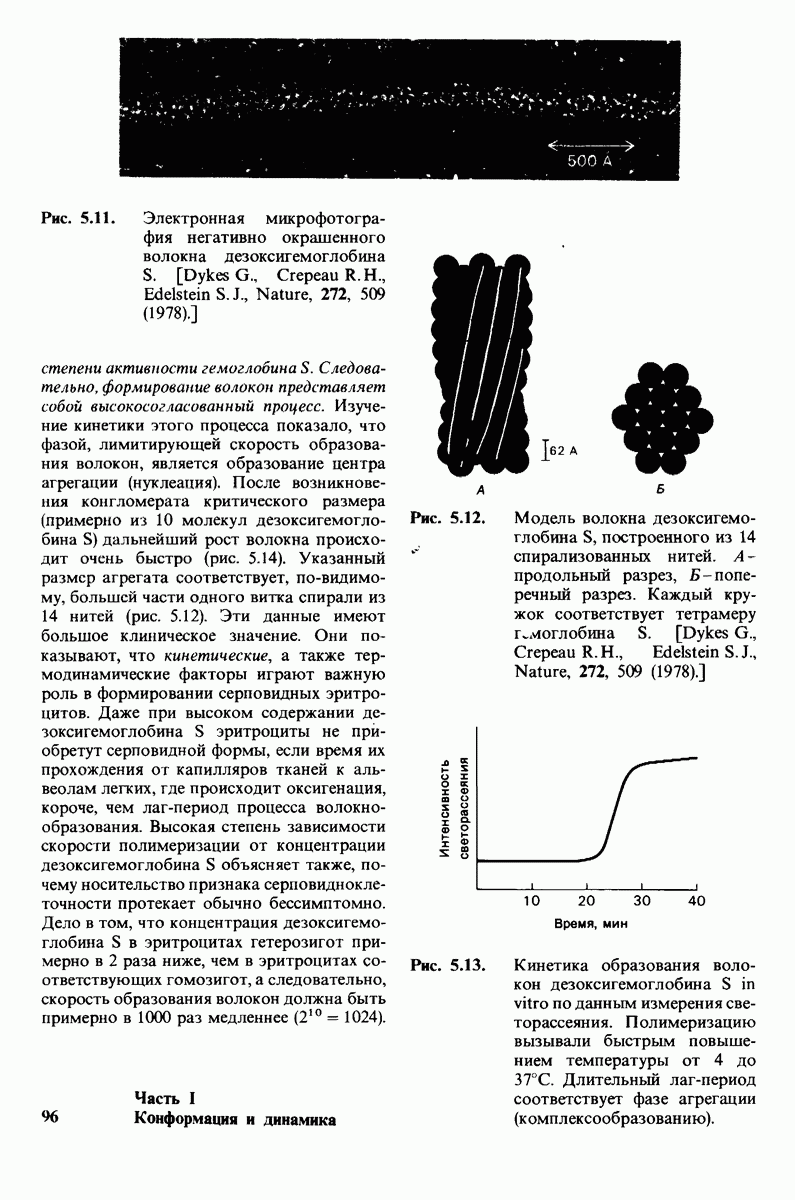
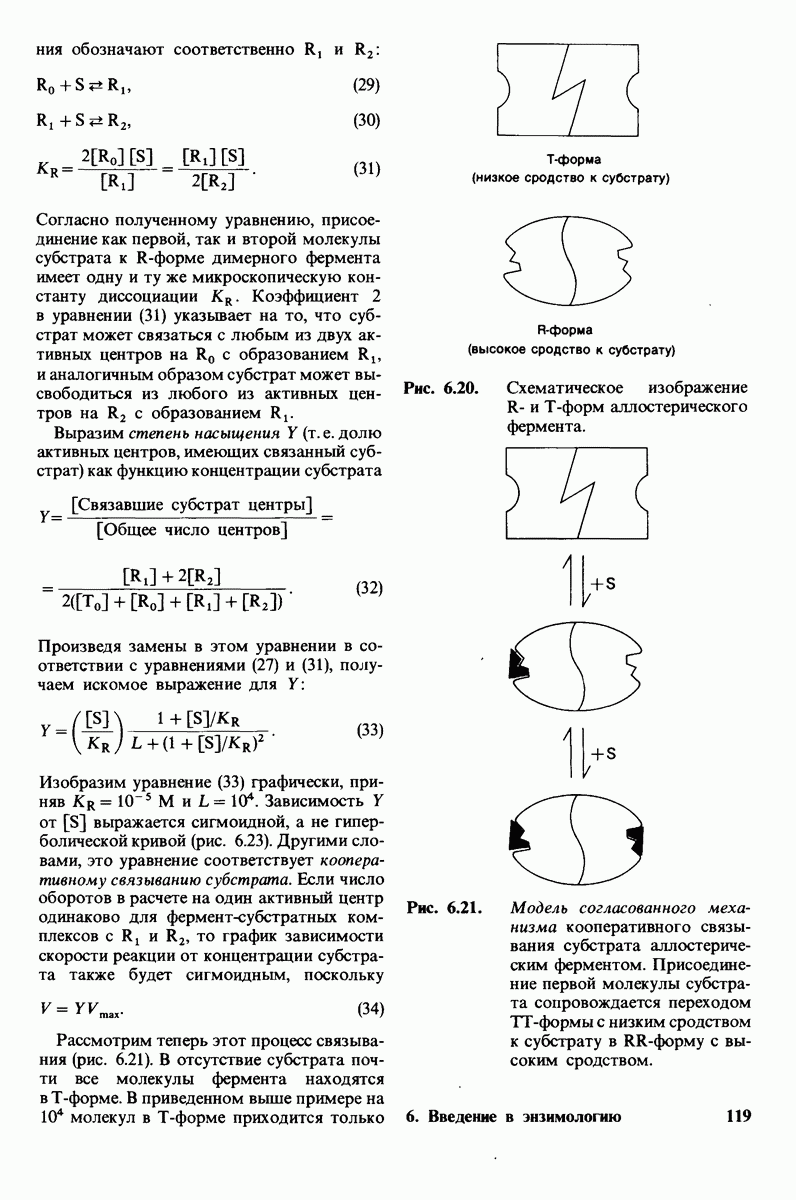
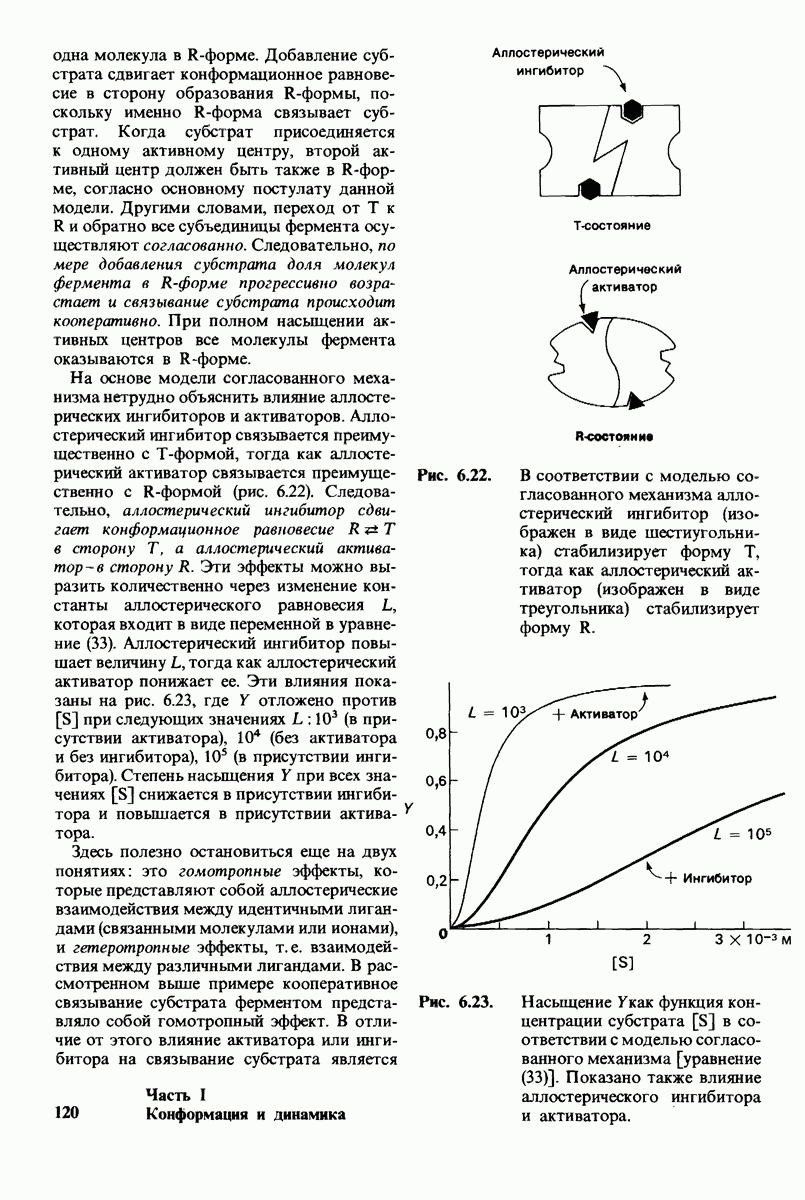
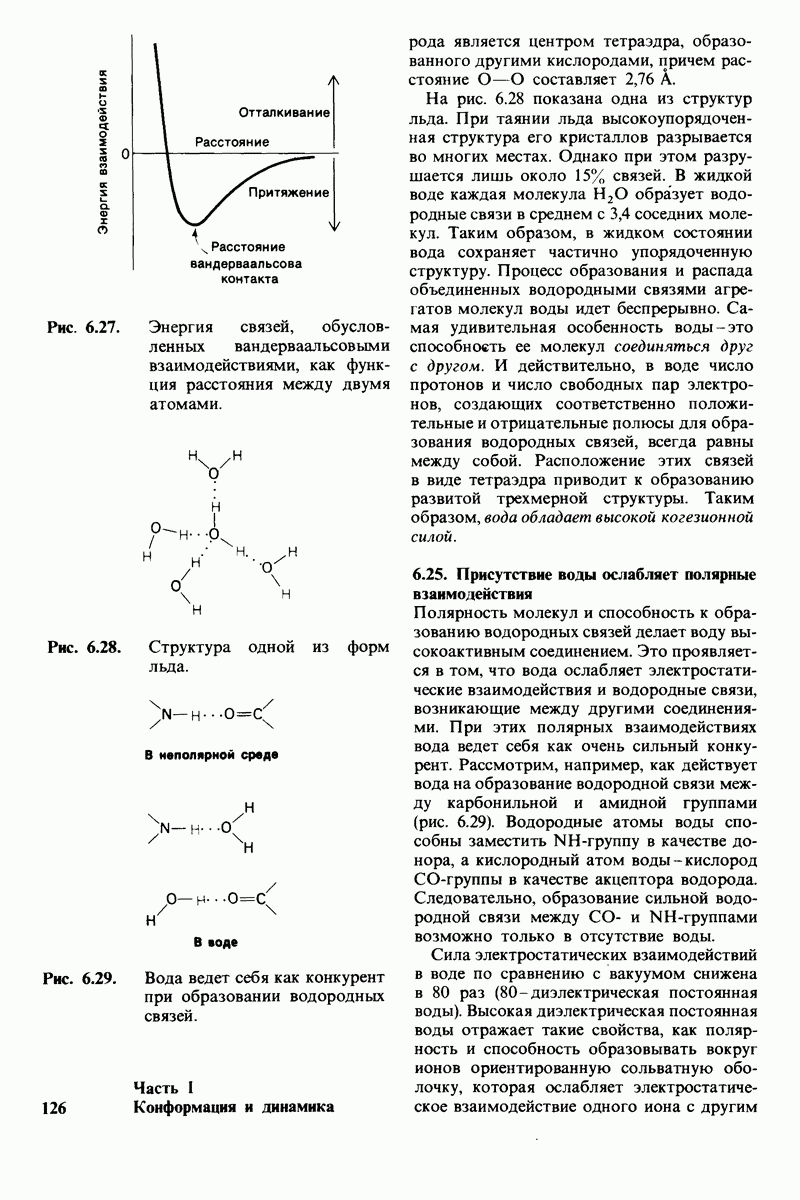
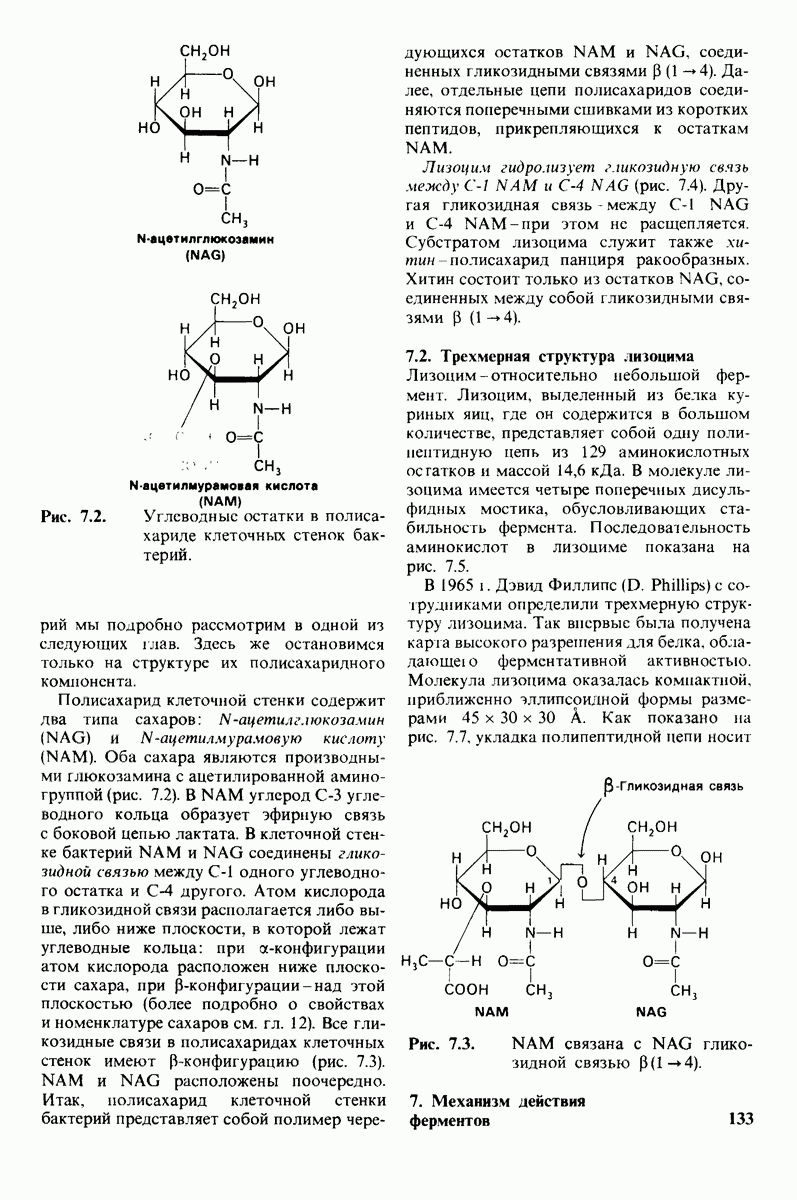
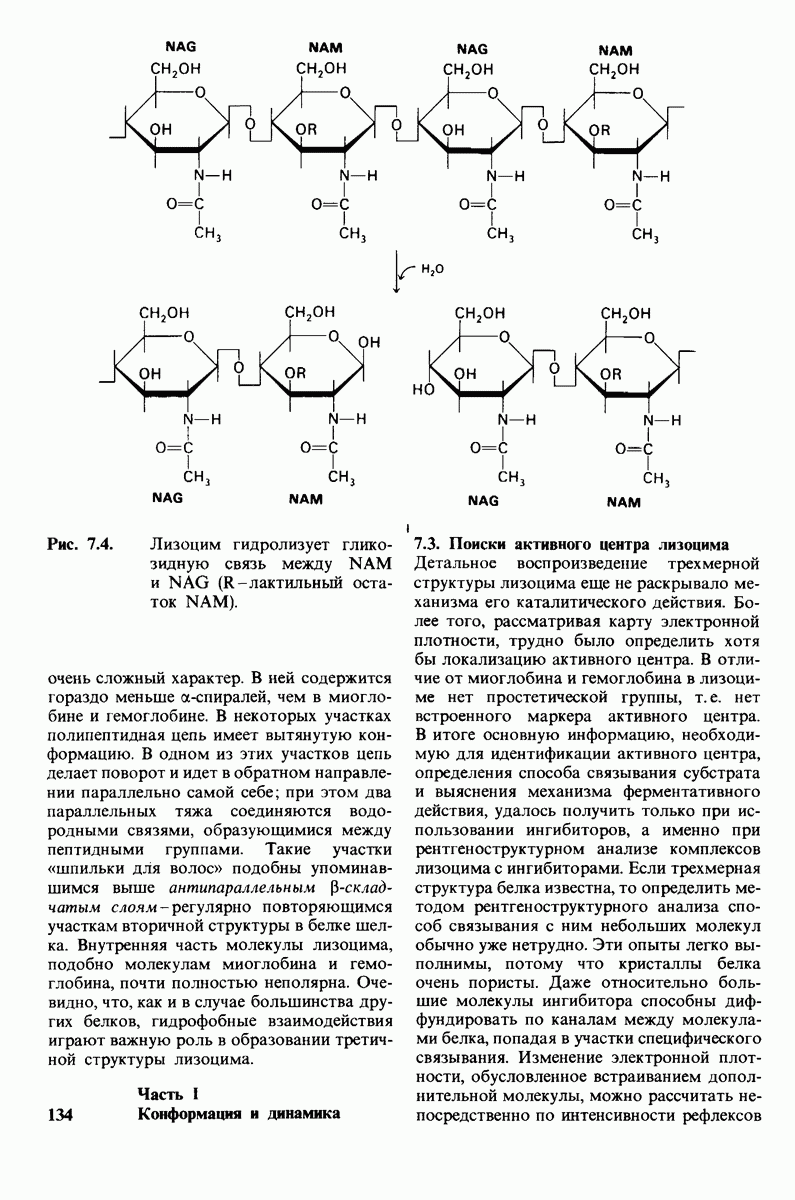
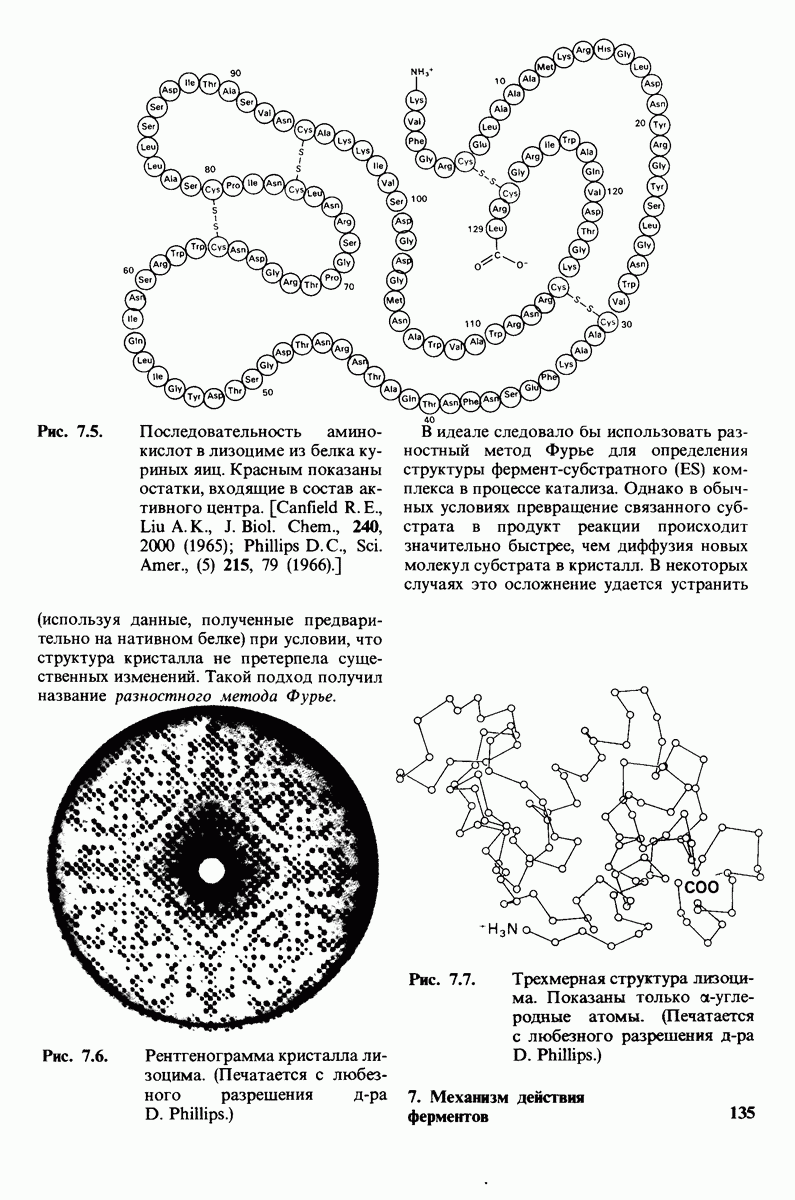
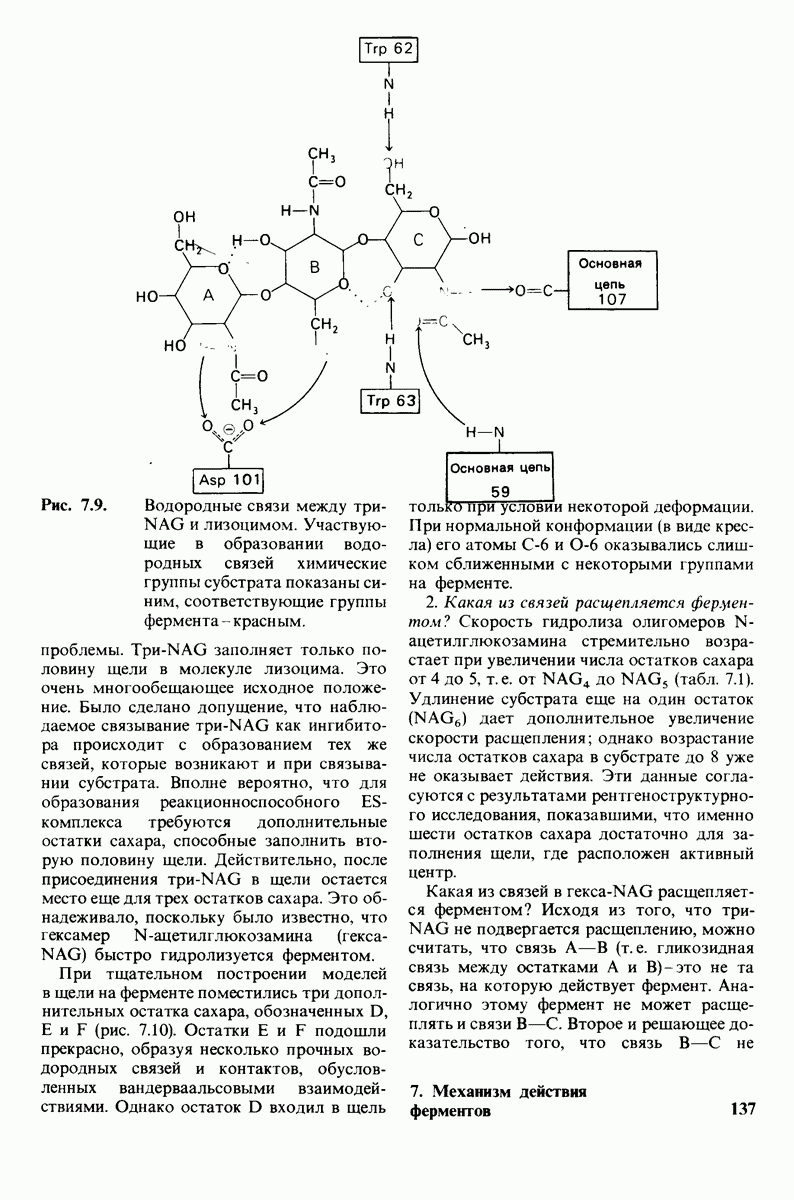
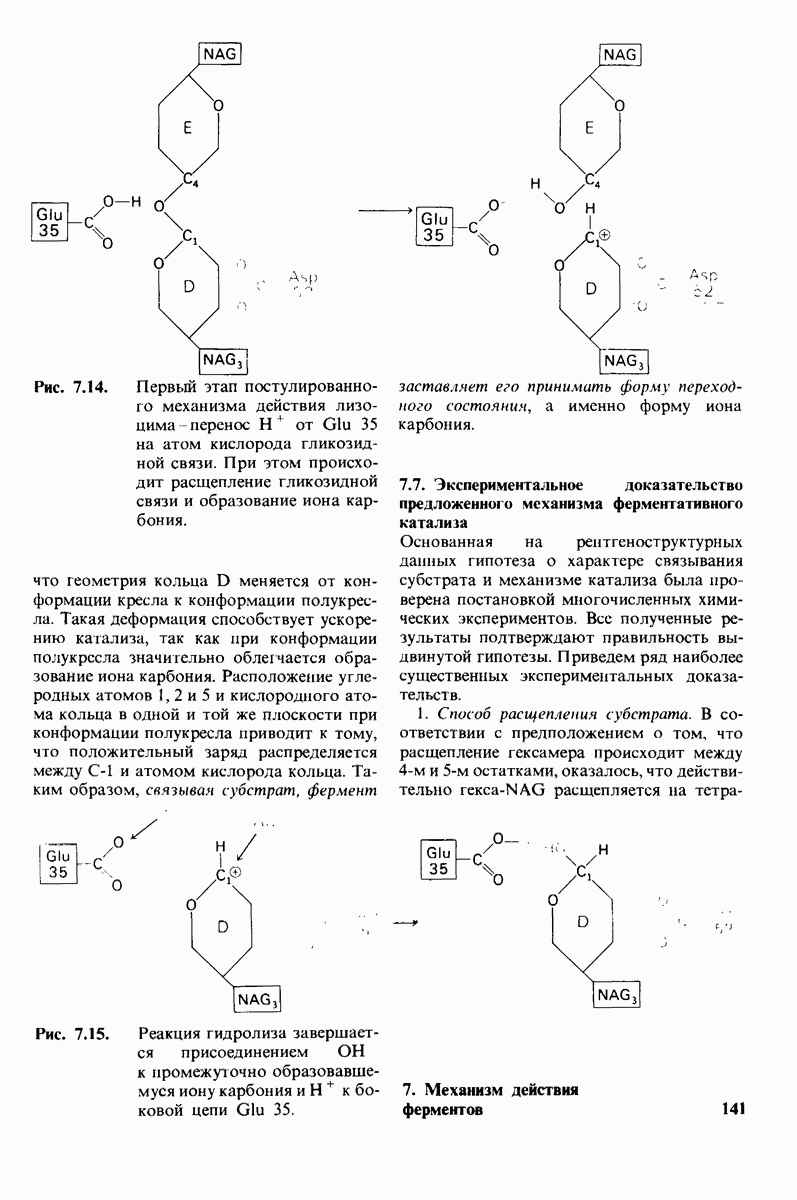
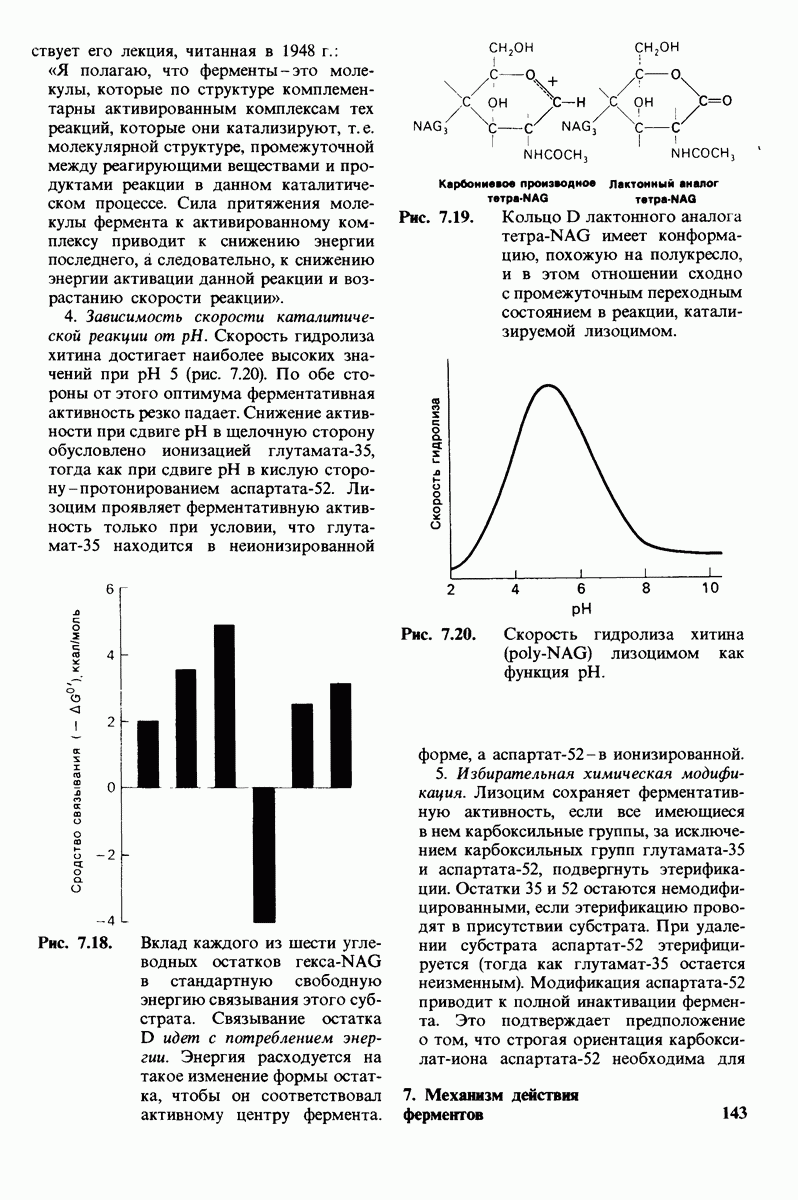
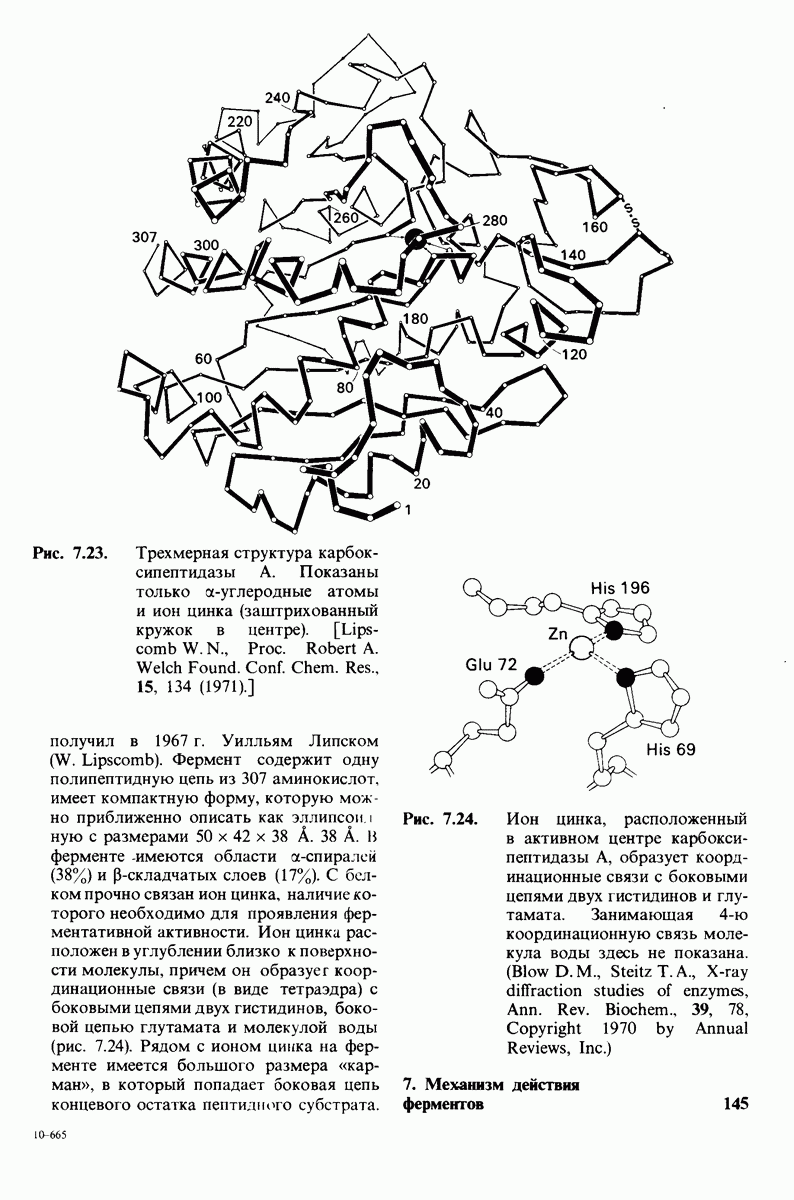
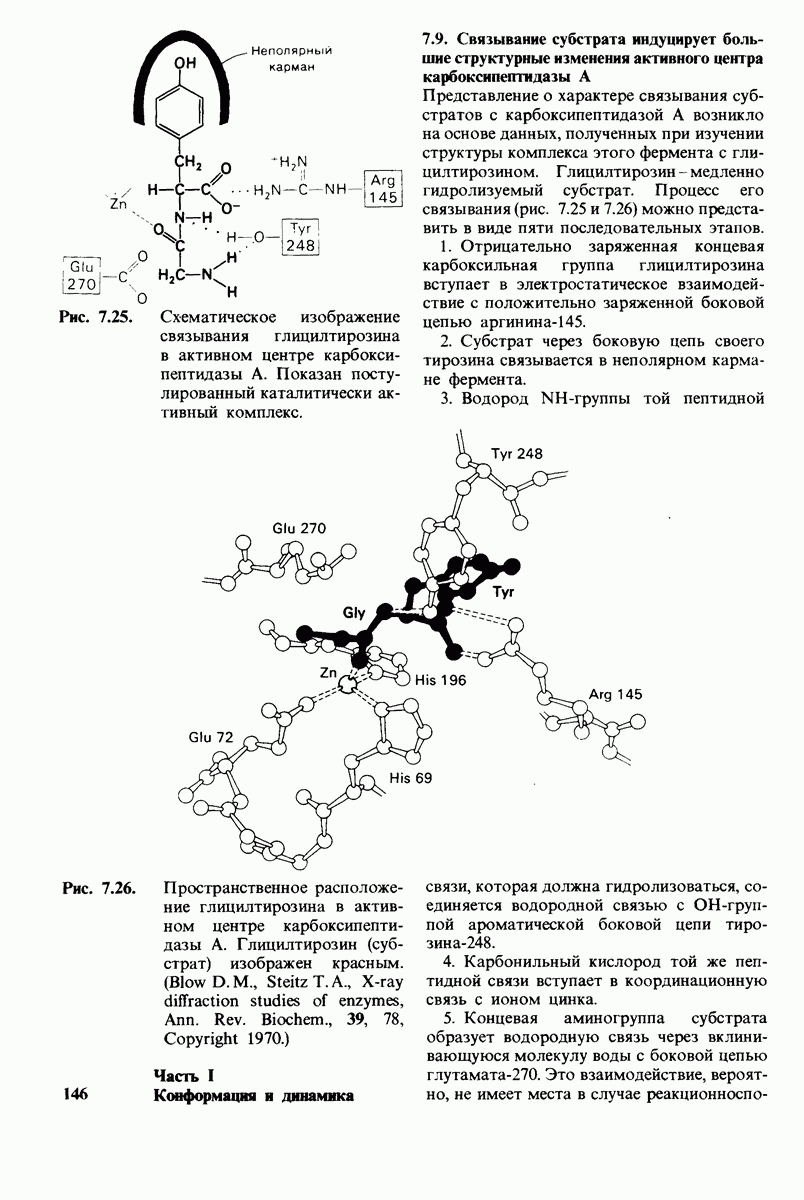
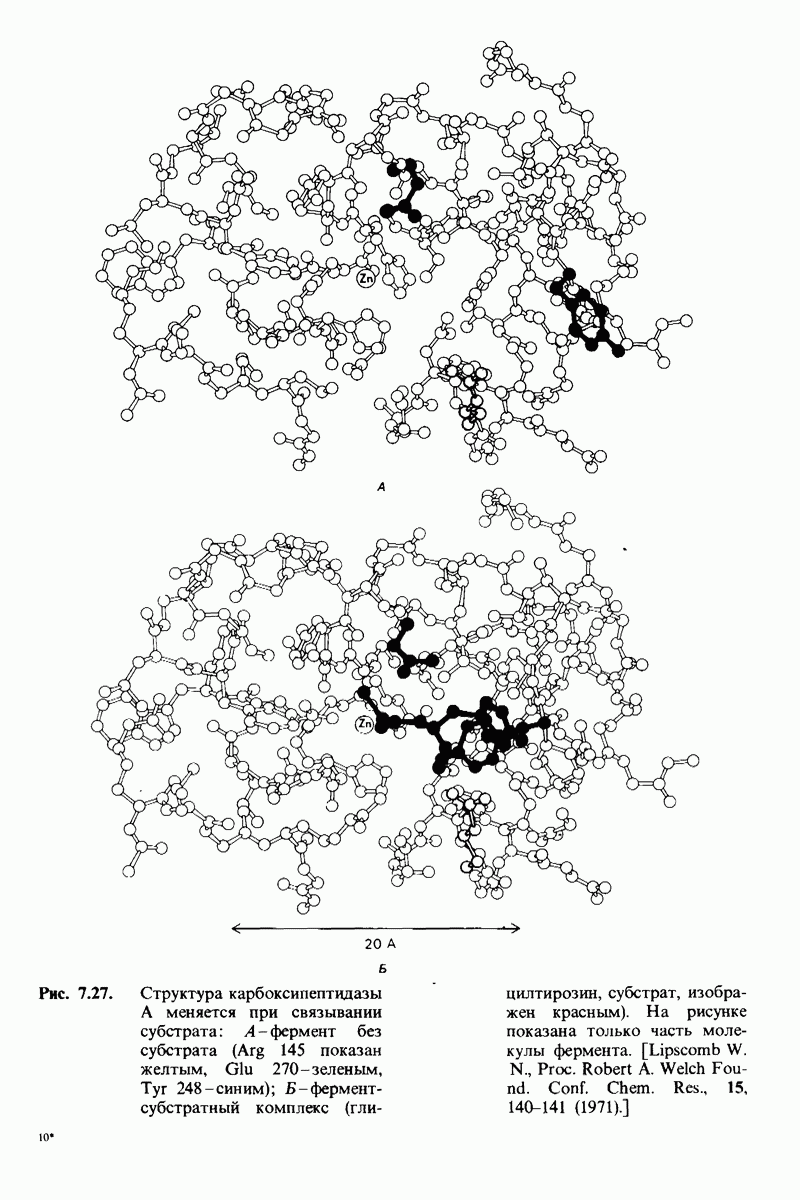
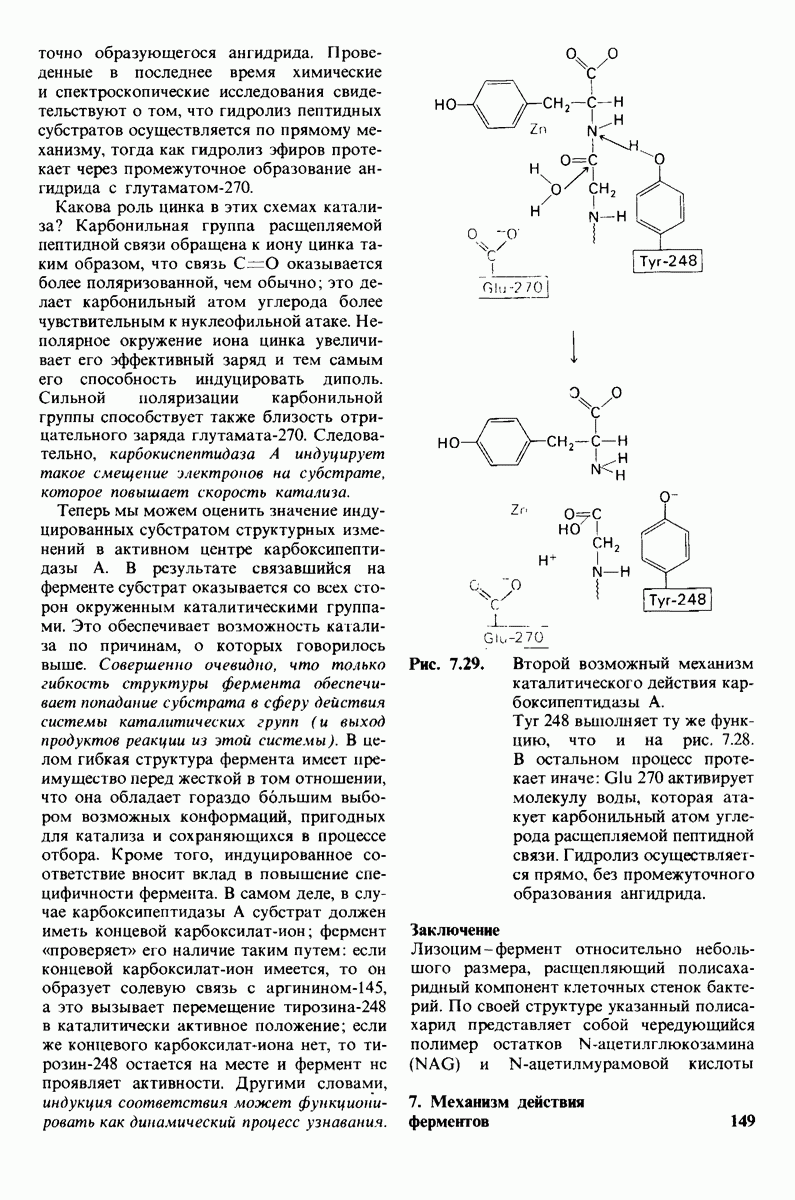
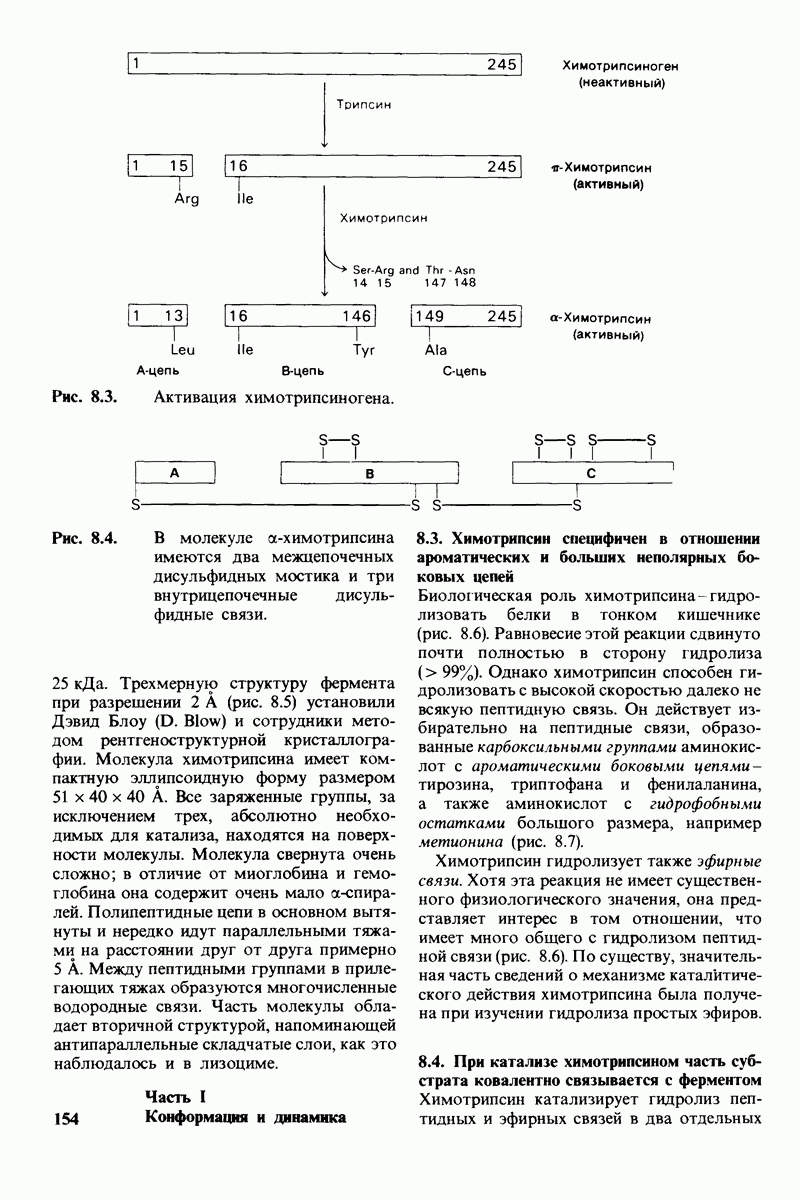
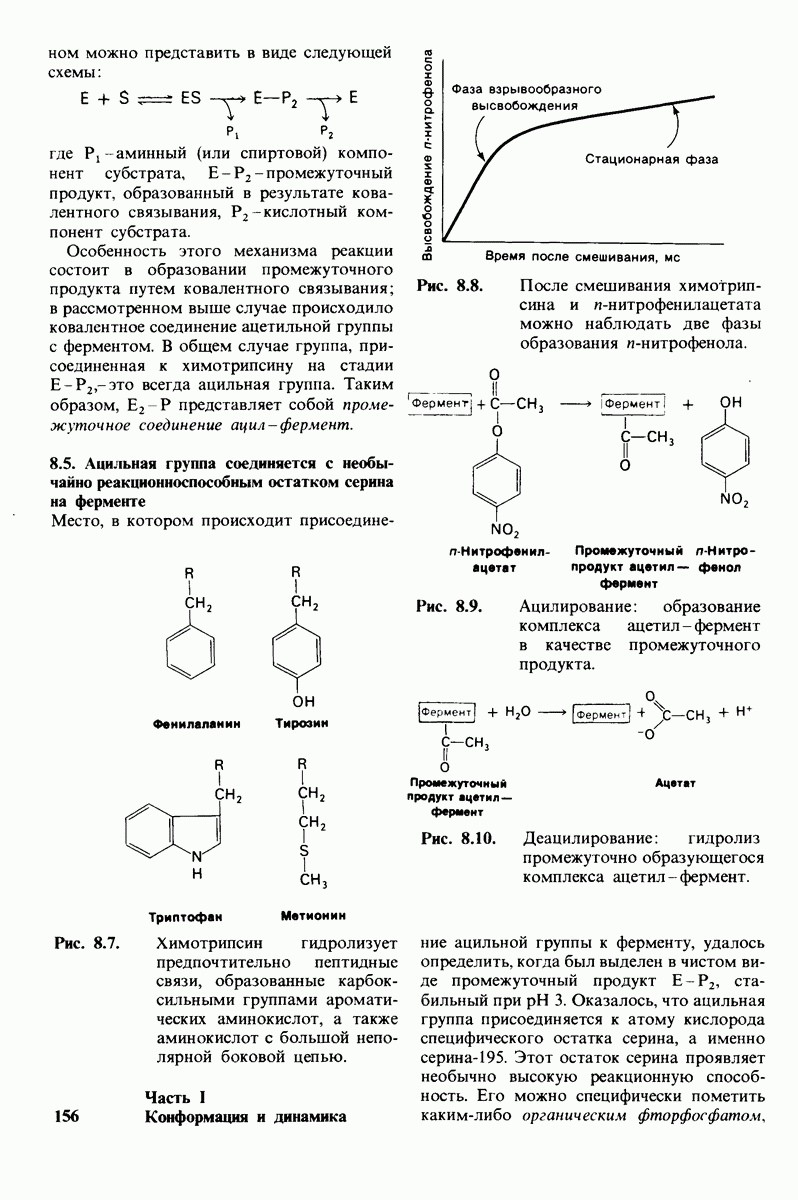
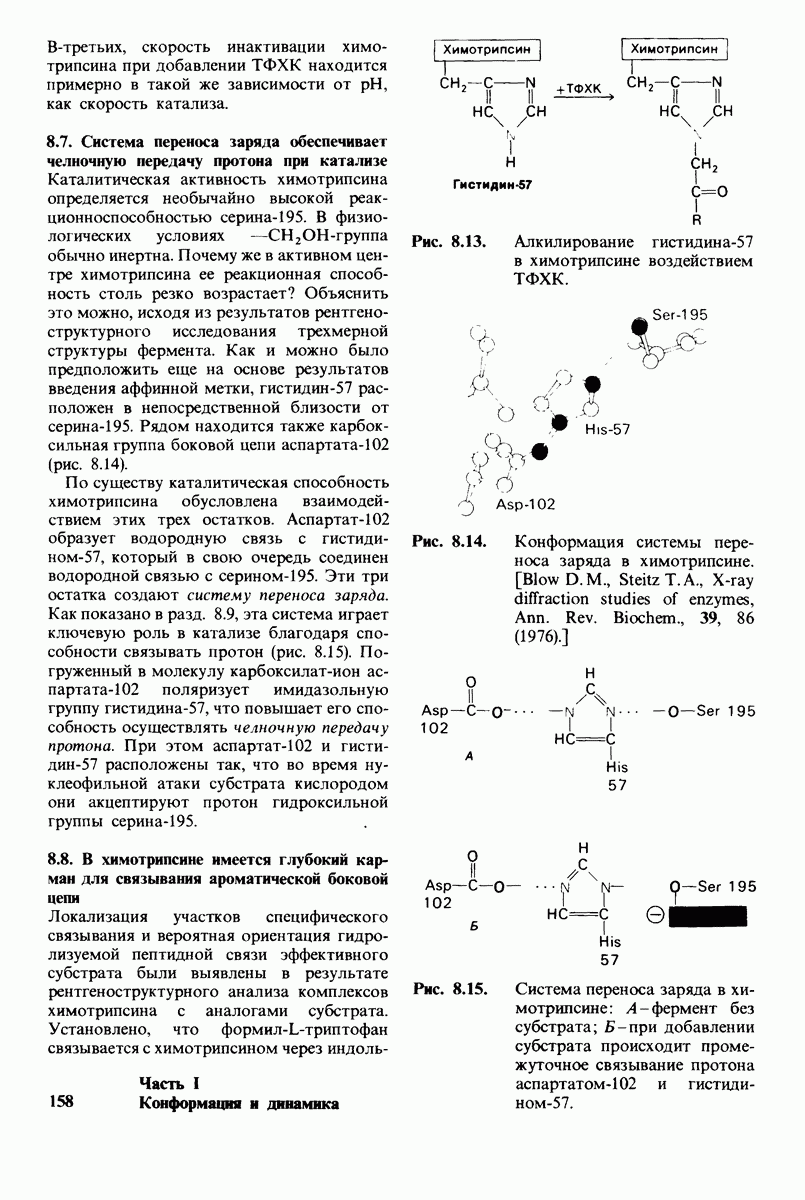
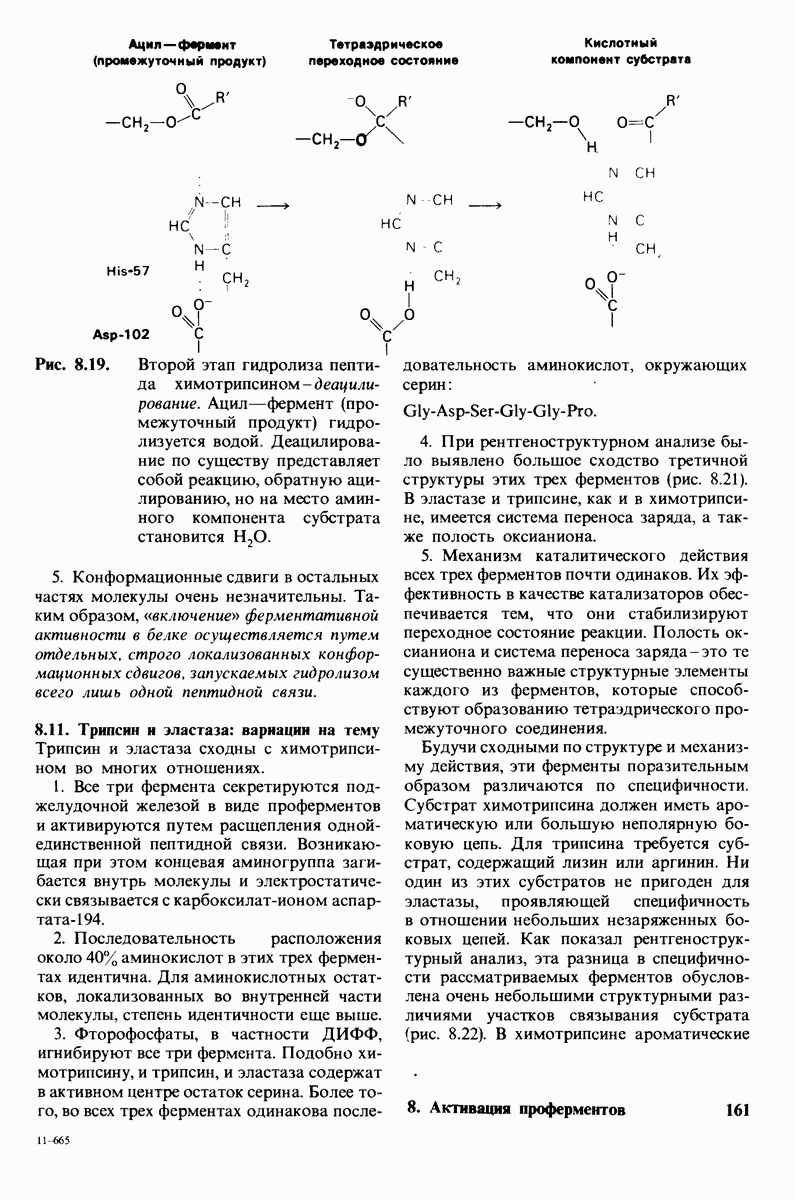
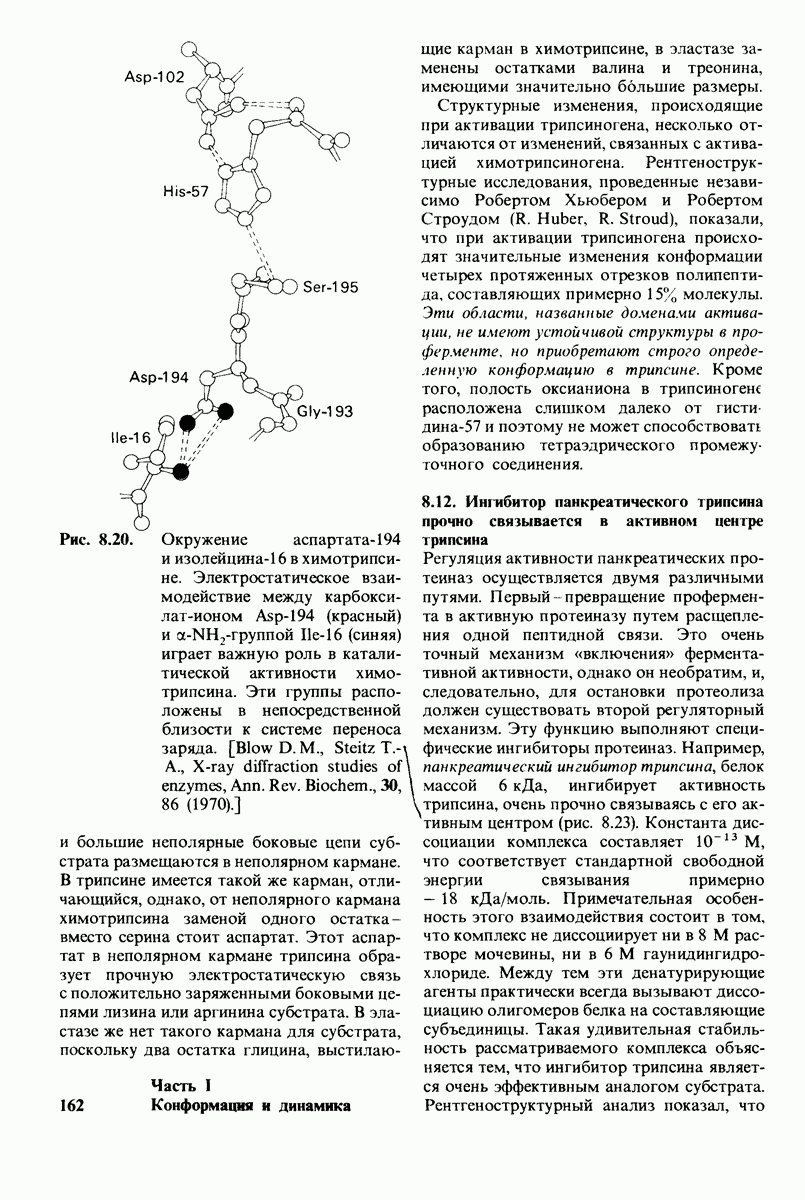
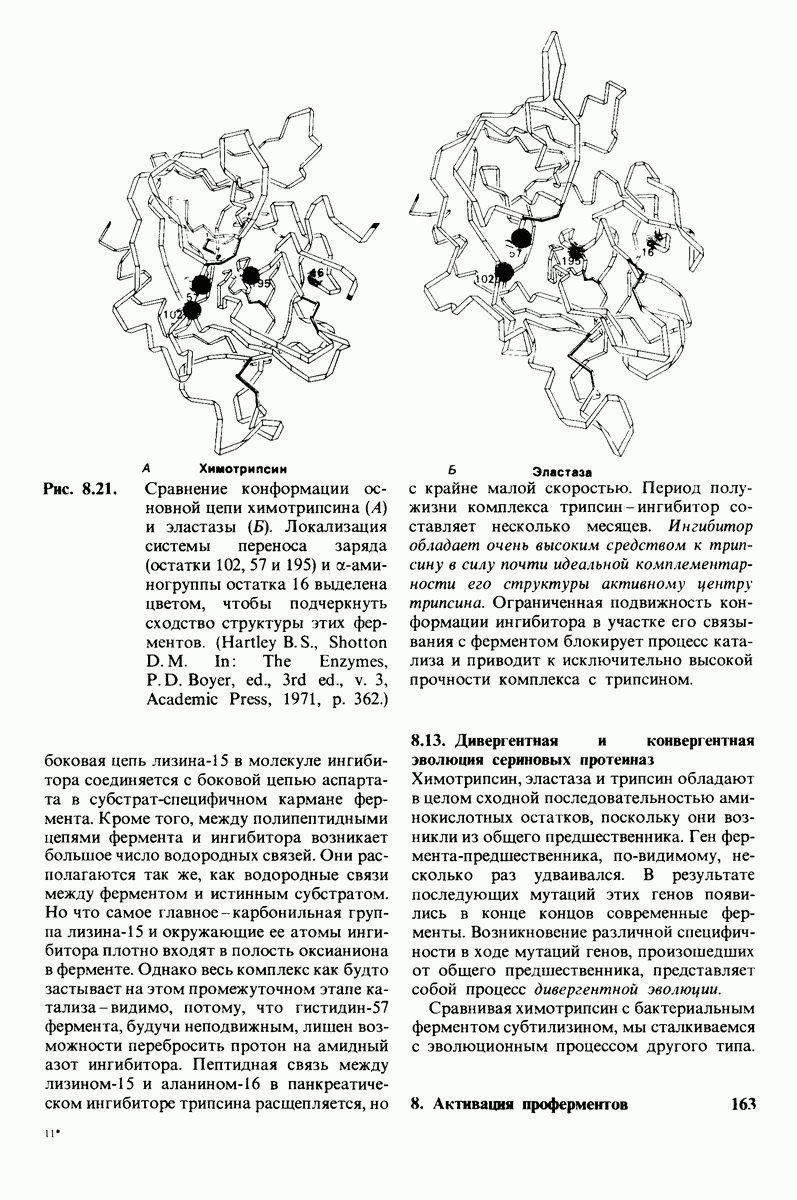
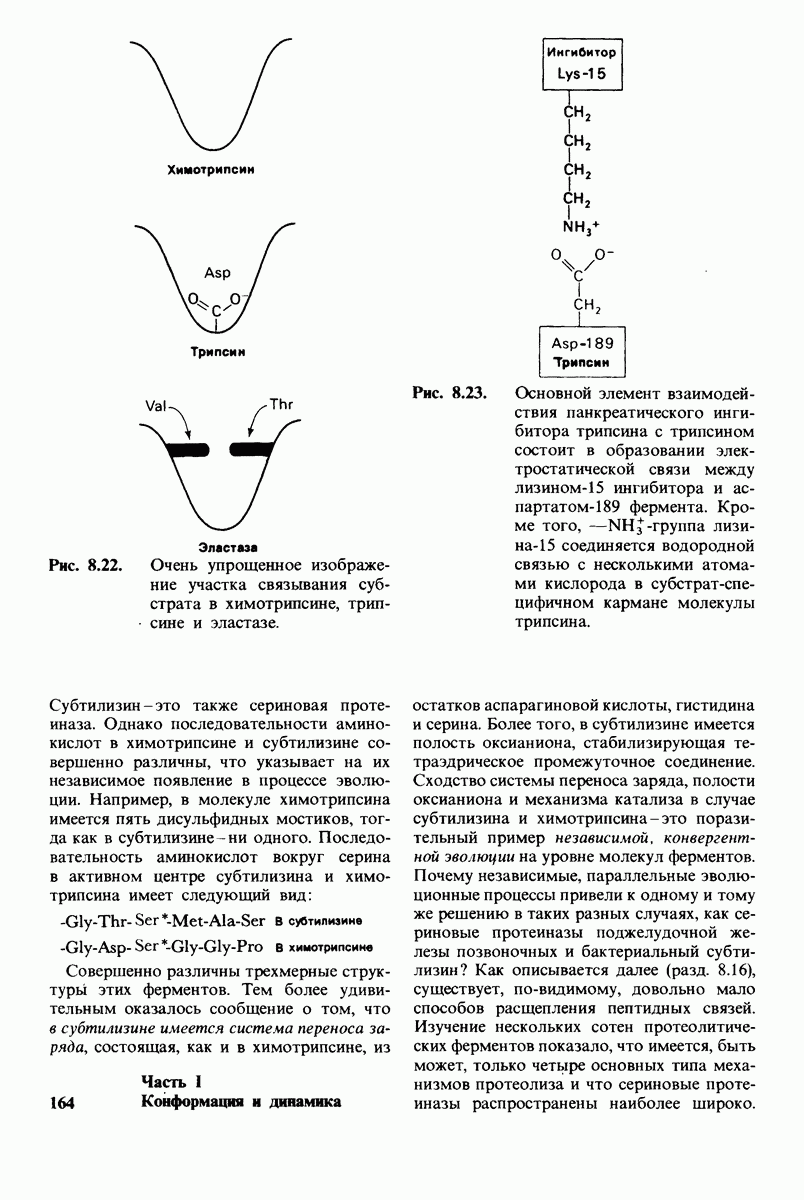
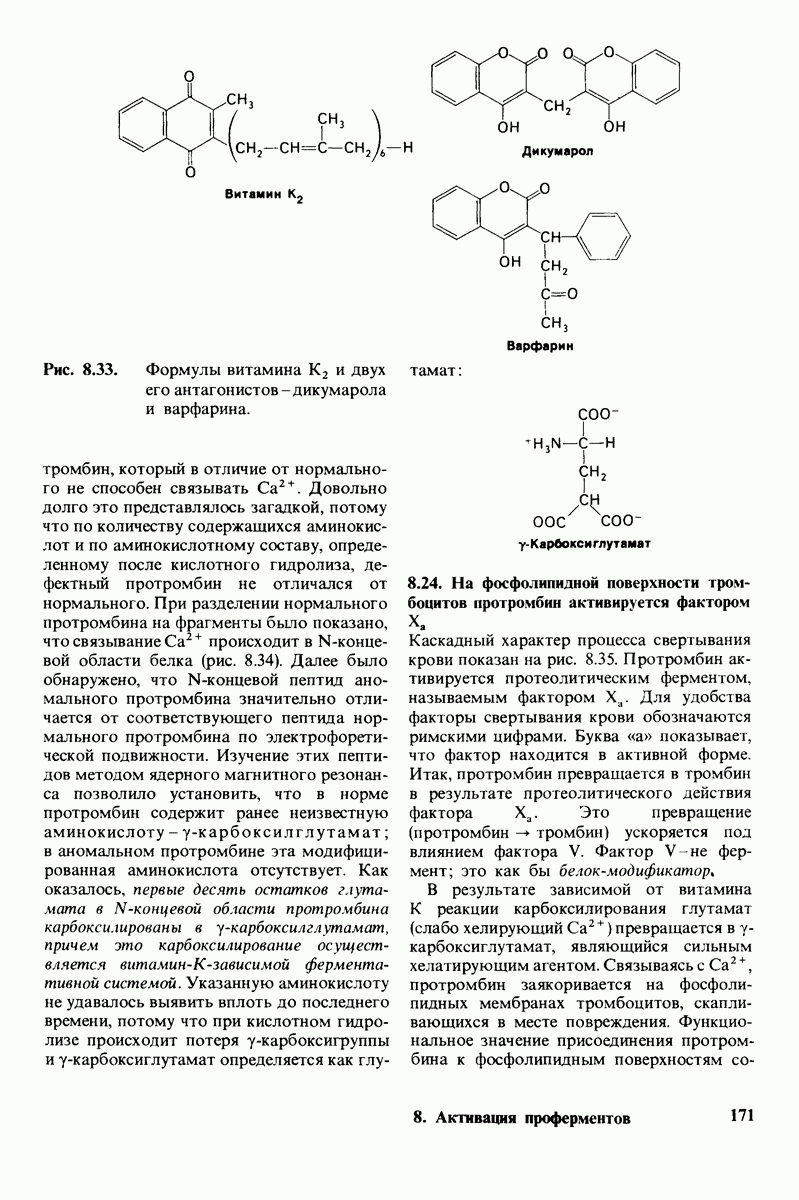
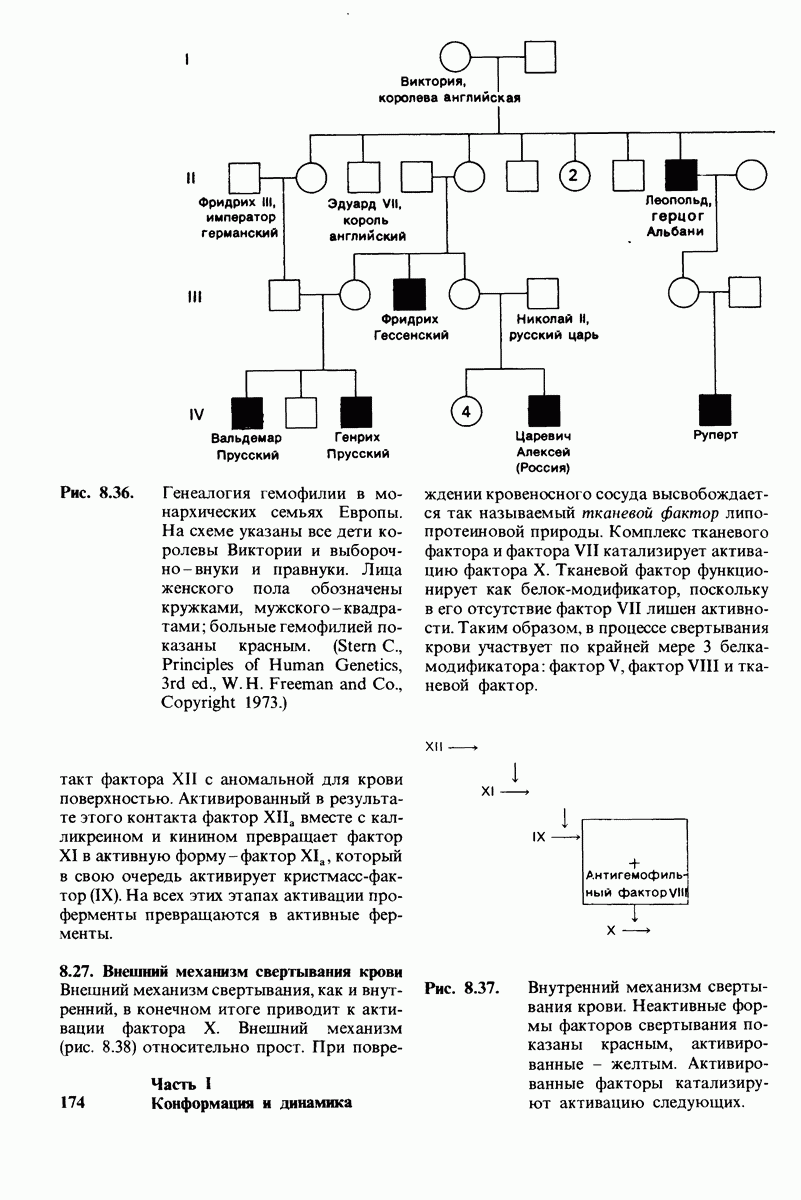
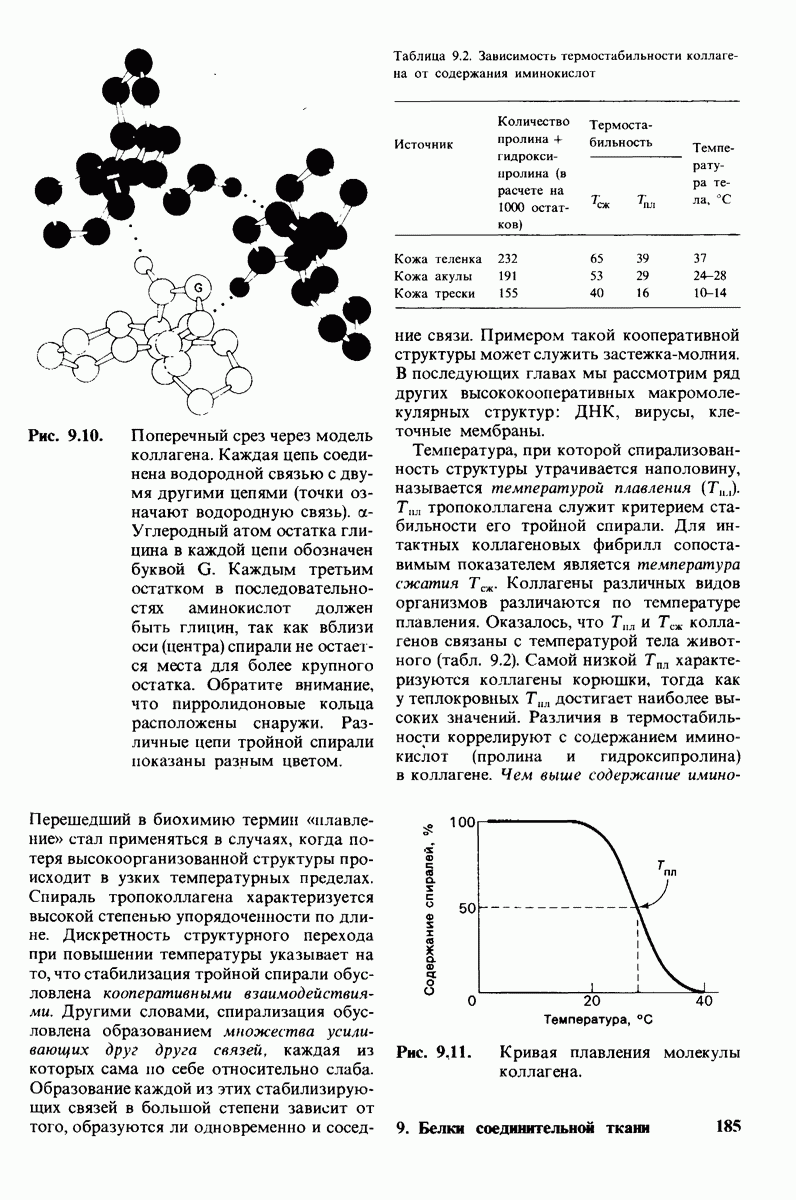
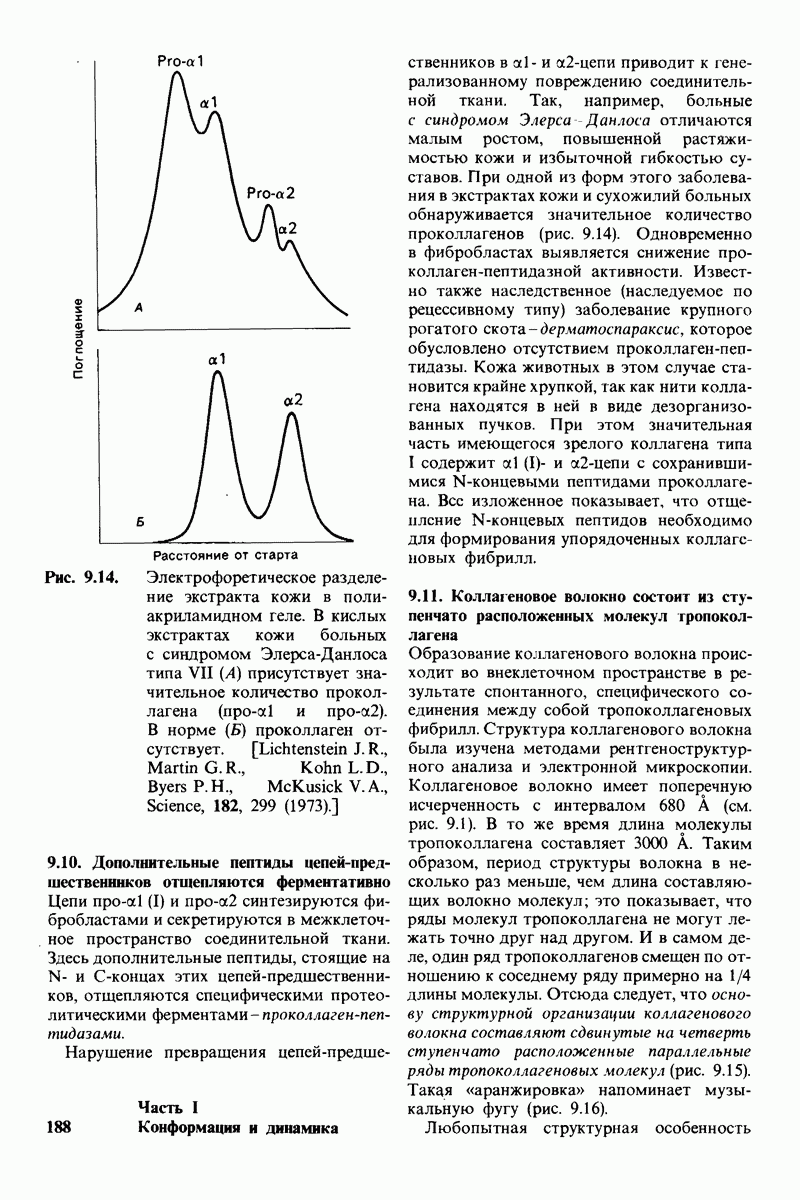
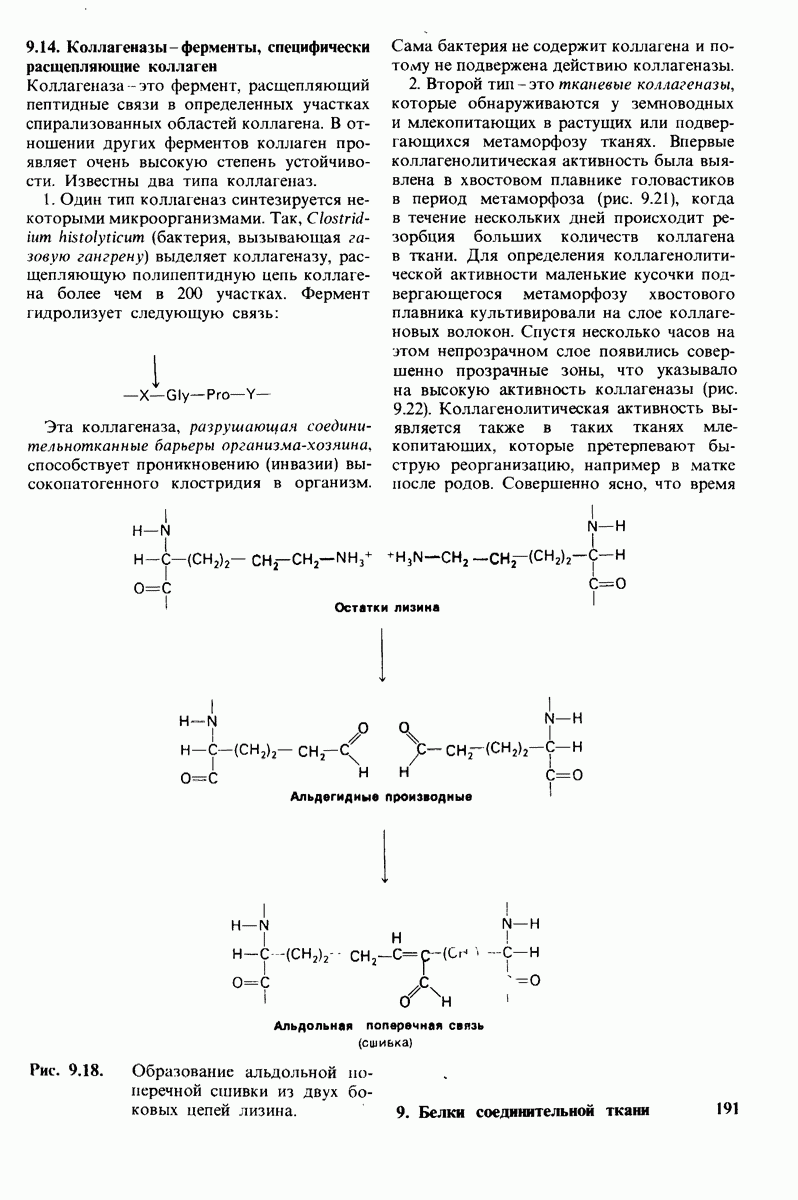
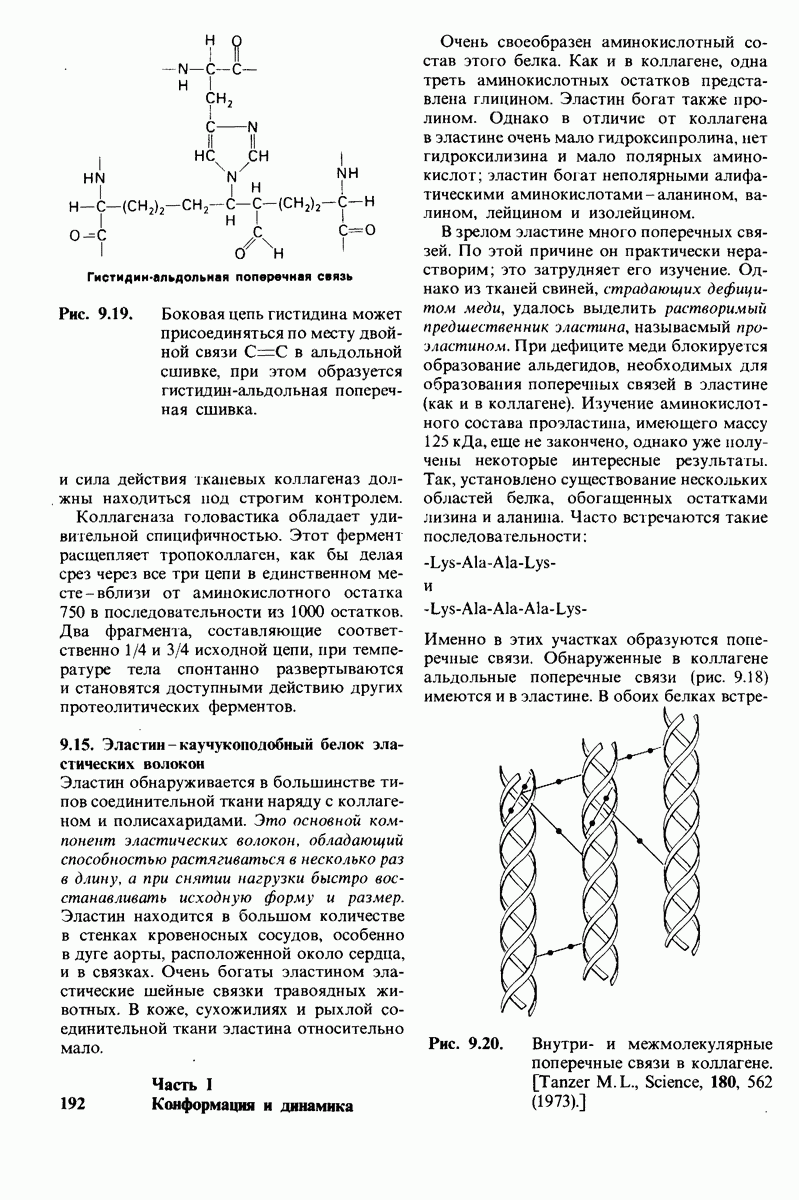
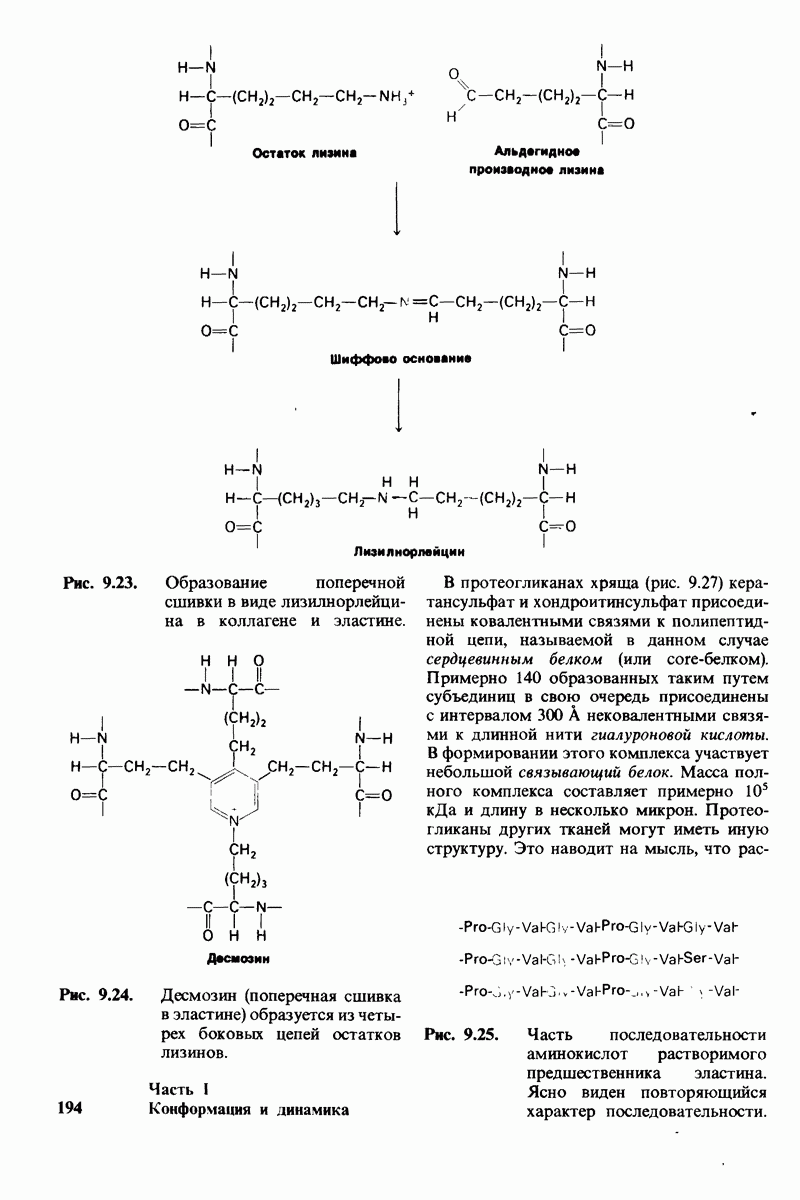
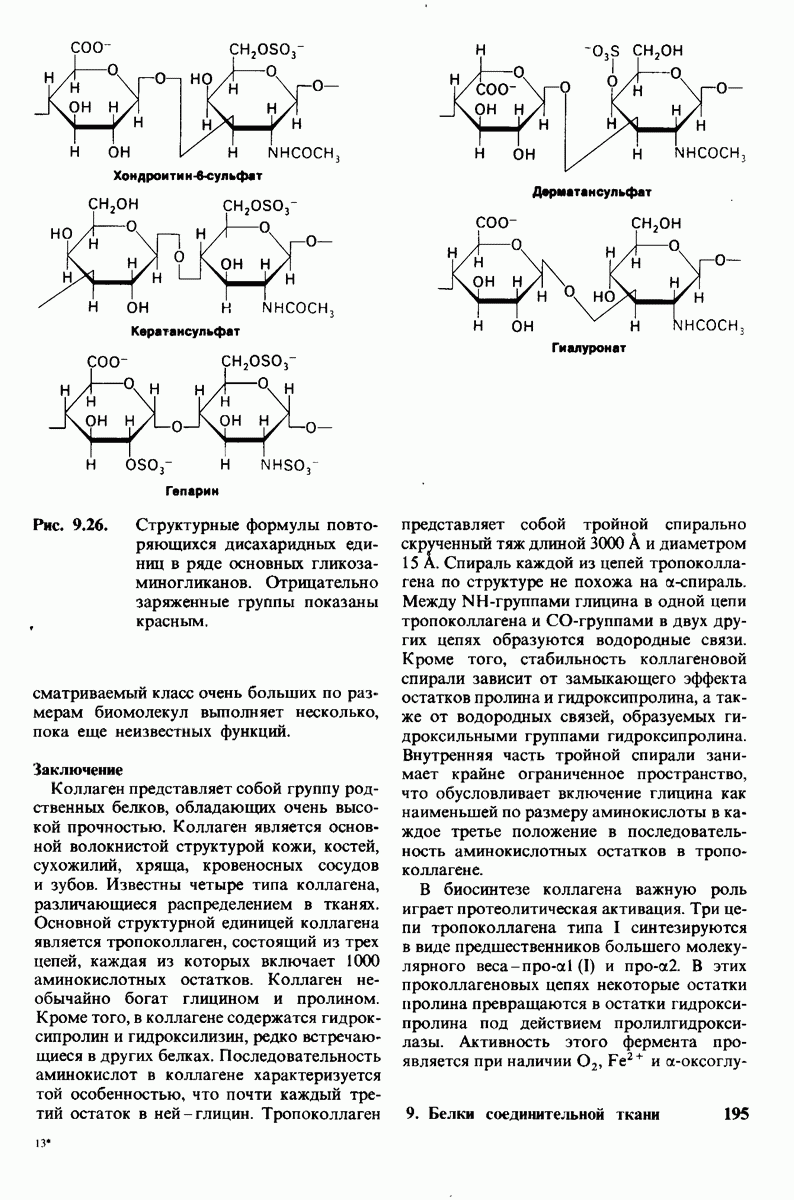
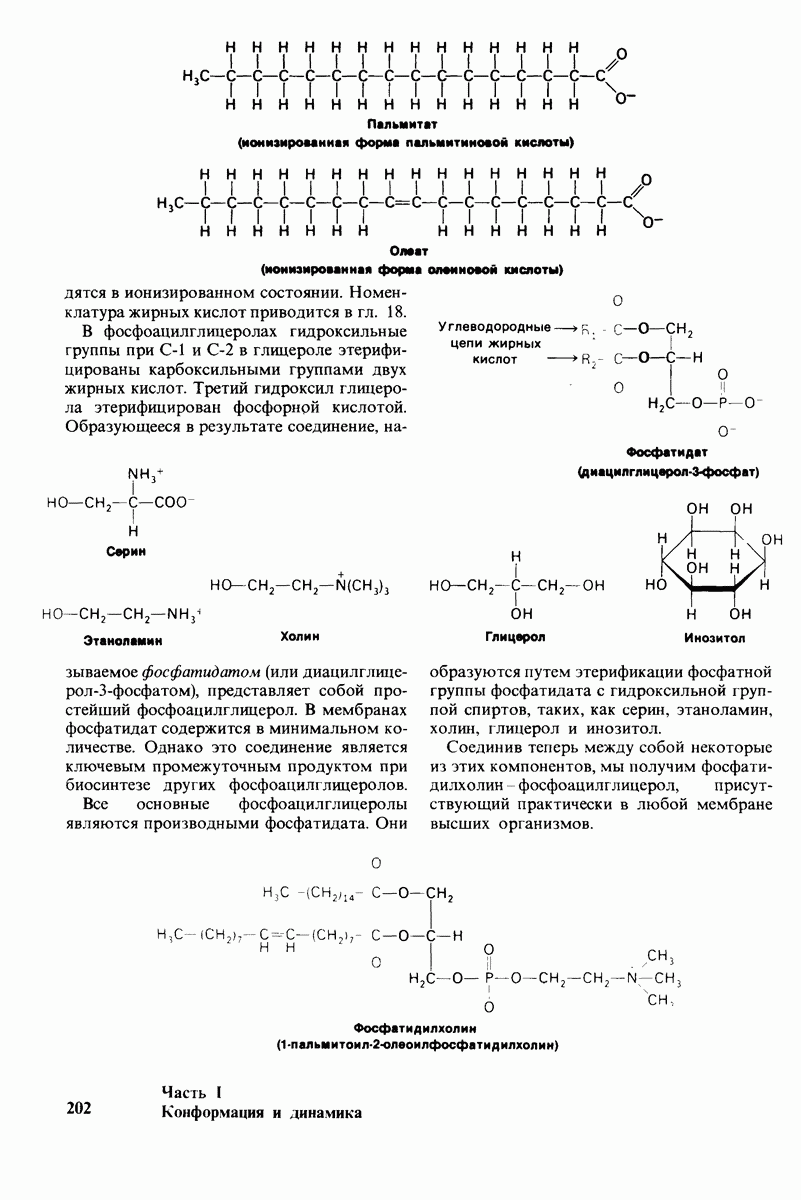
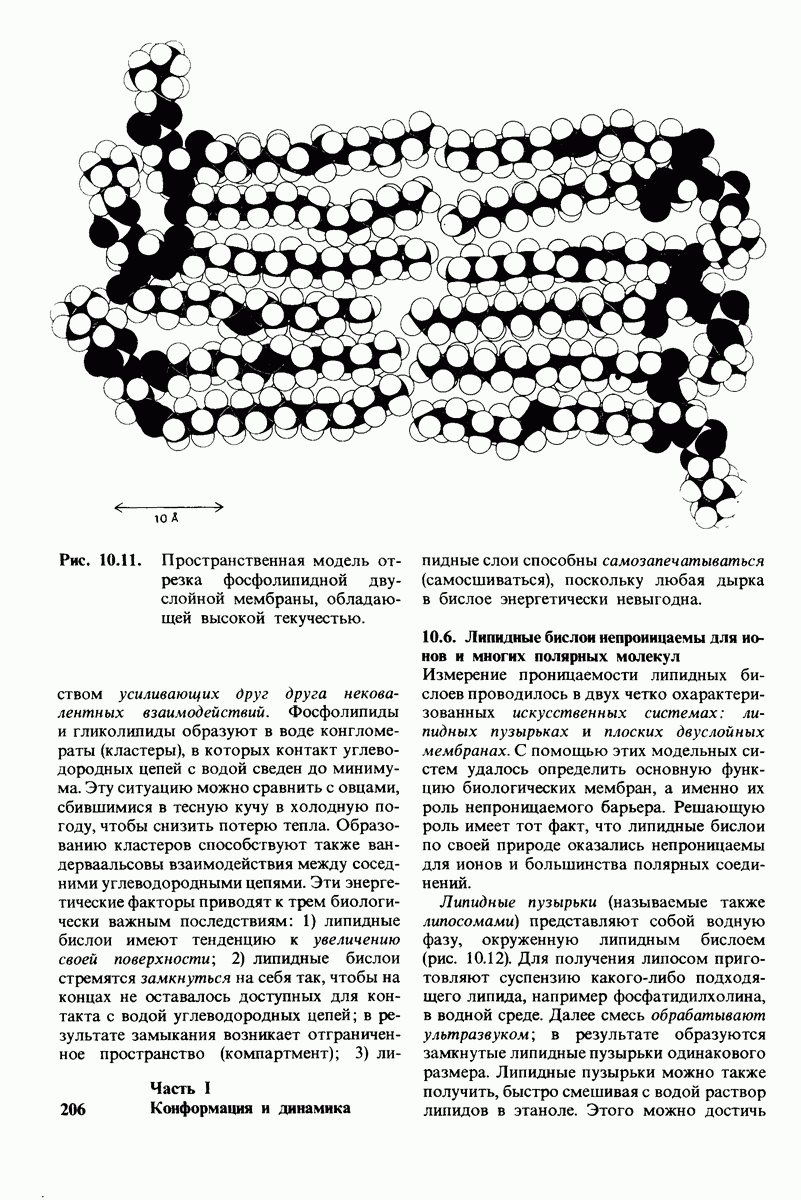
10.12. Углеводные единицы расположены на наружной стороне плазматической мембраны
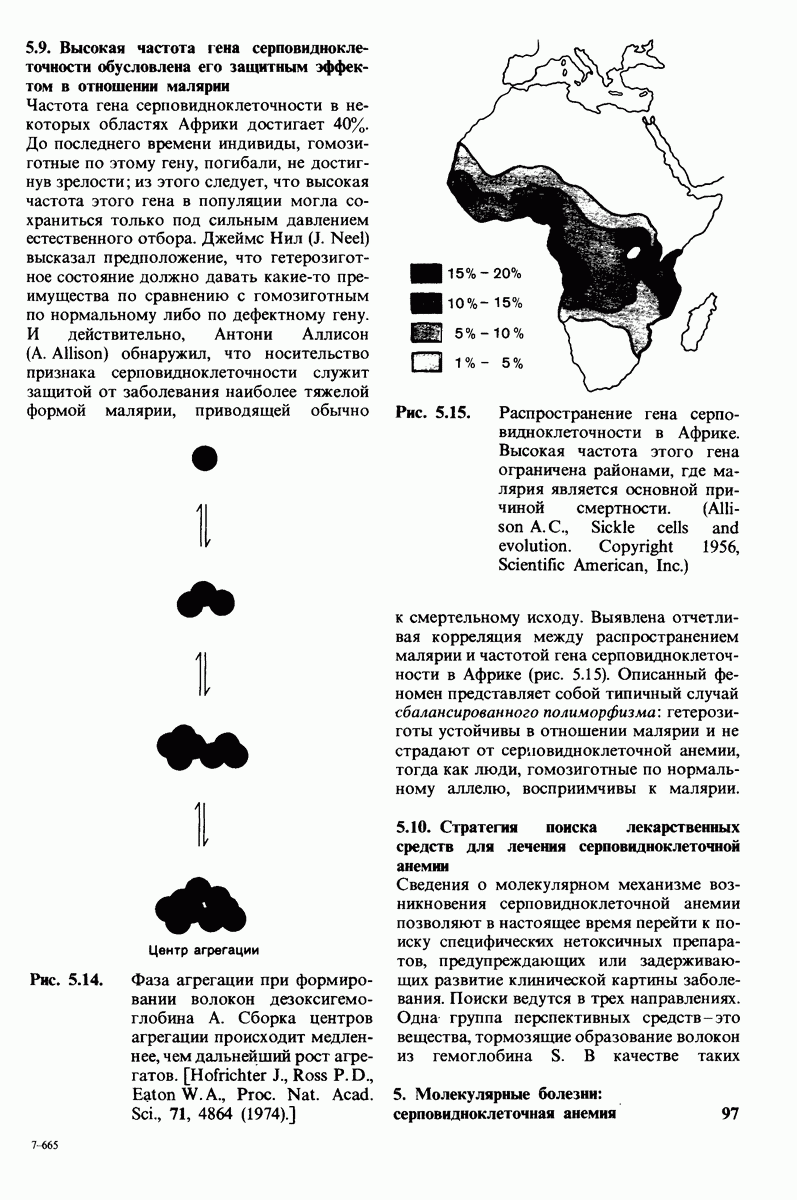
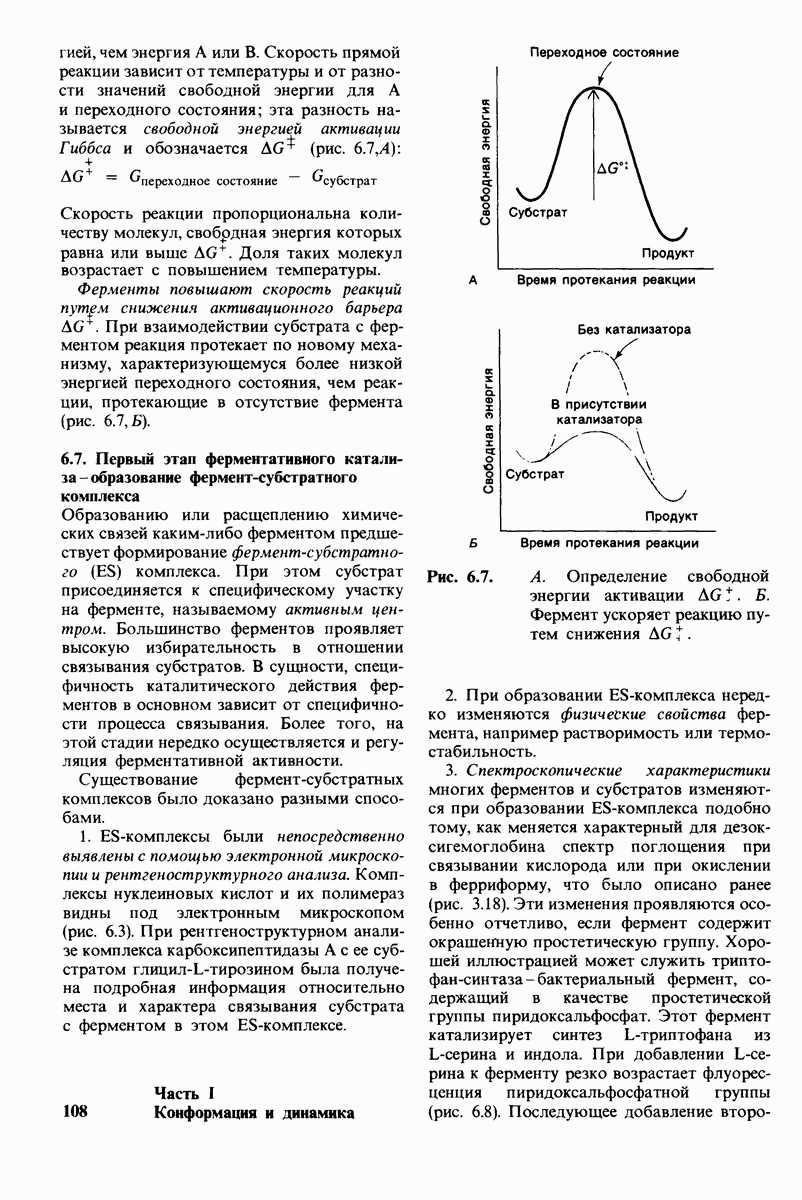
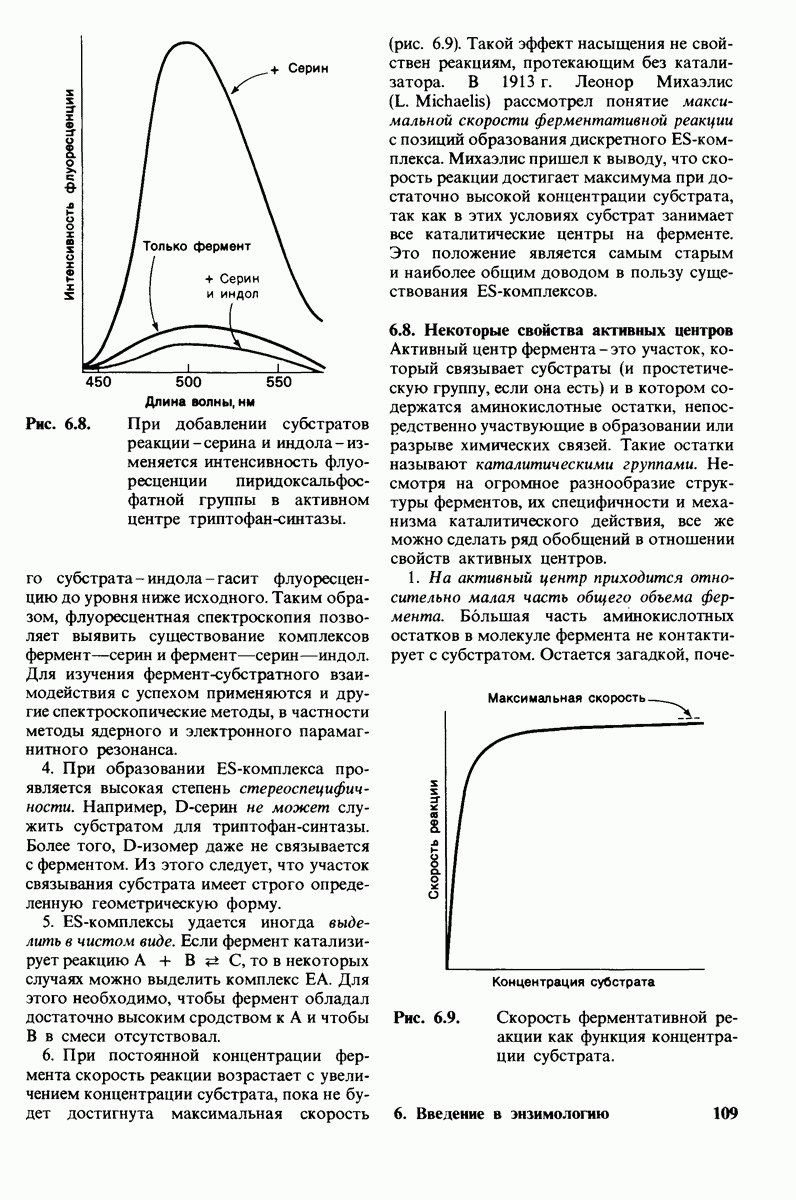
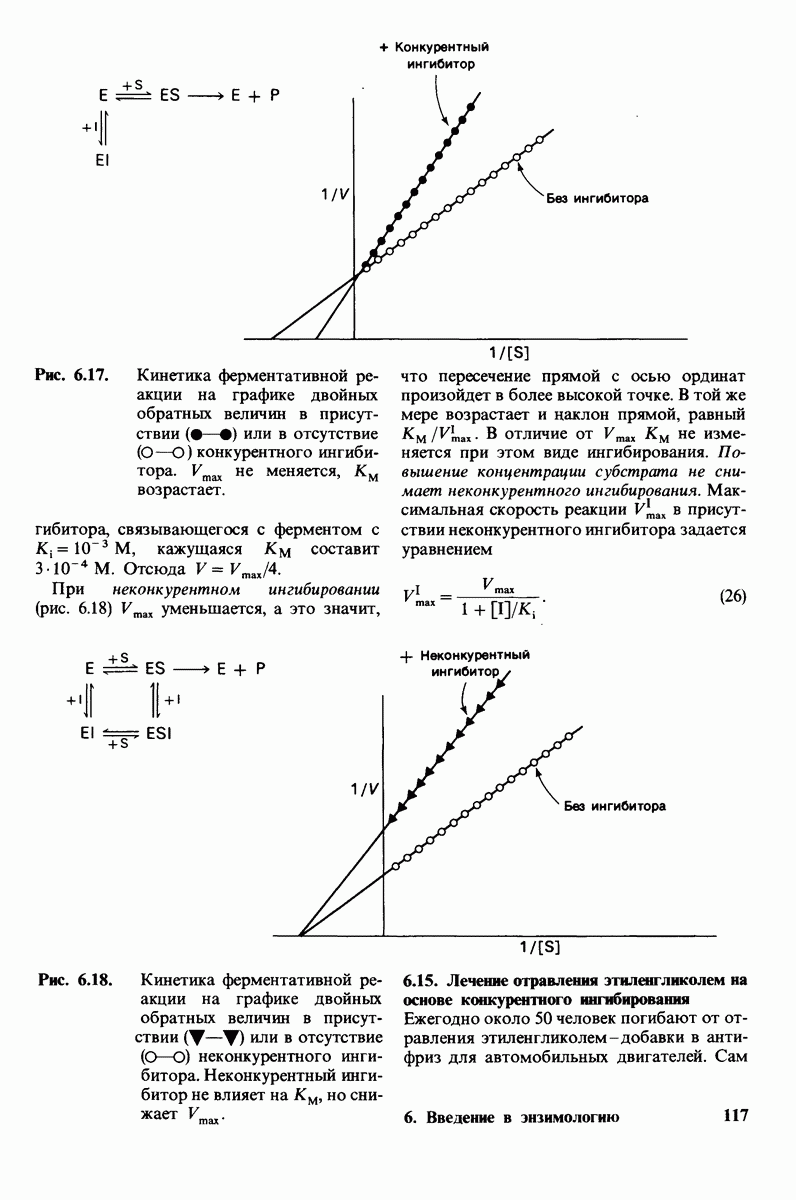
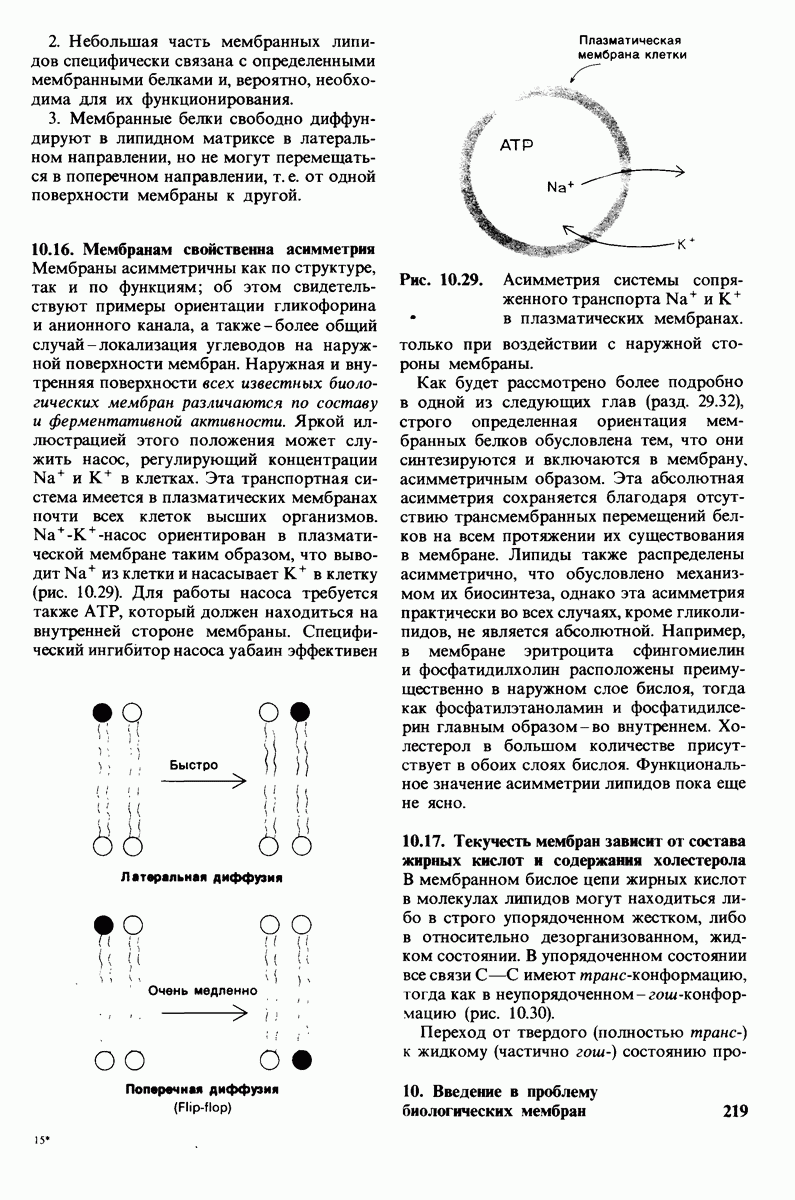
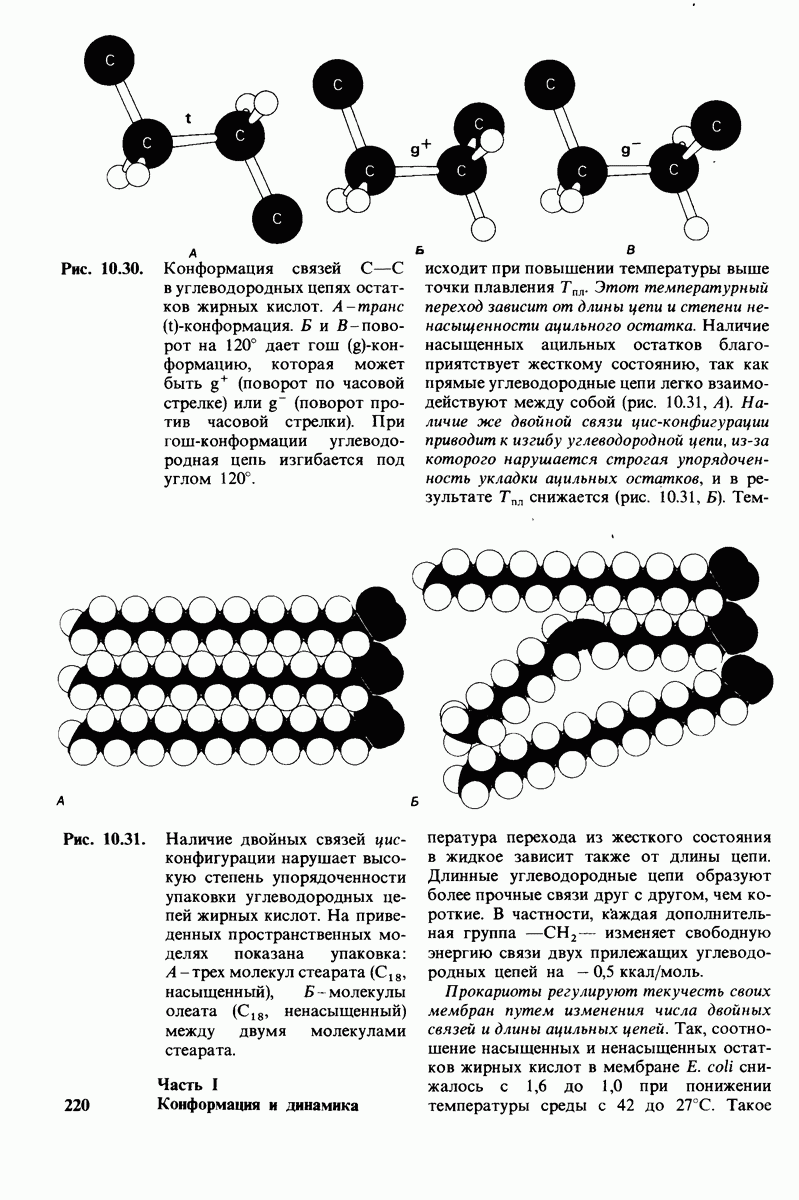
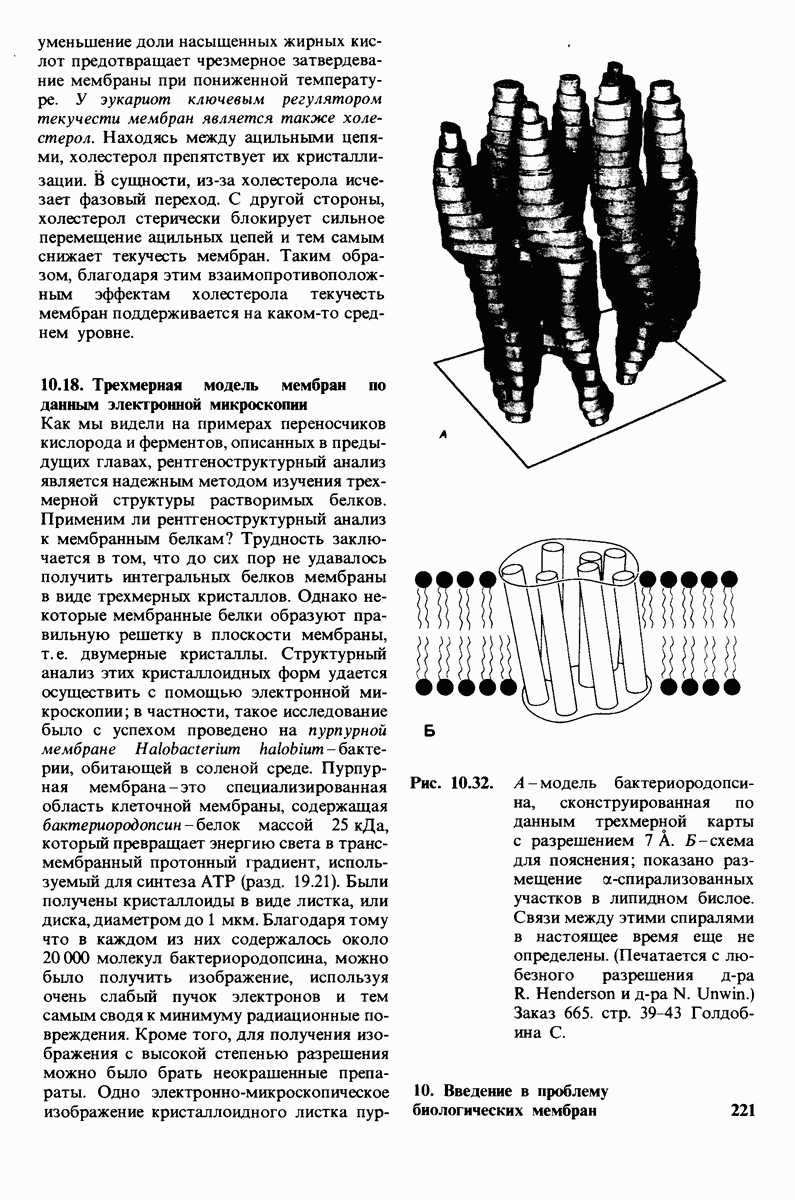
В клеточных мембранах эукариот содержится от 2 до 10% углеводов в форме гликолипидов и гликопротеинов. Как было сказано выше (разд. 10.3), гликолипиды высших организмов представлены производными сфингозина с одним или более остатками сахара. В мембранных гликопротеинах одна или несколько углеводных цепей присоединены к боковым цепям серина, треонина или аспарагина (обычно через N-ацетилглюкозамин или N-ацетилгалактозамин). Локализацию этих углеводных групп в мембране определяют путем использования специфической метки. Подходящей меткой оказались лектины - растительные белки, обладающие высоким сродством к специфическим углеводным остаткам. Например, конканавалин А присоединяется к внутренним и нередуцирующим концевым остаткам  -маннозы, тогда как агглютинин из зародыша пшеницы присоединяется к концевым остаткам N-ацетилглюкозамина. Эти лектины хорошо видны в электронном микроскопе, когда они соединены с ферритином - белком, содержащим гидроксид железа и потому обладающим высокой электронной плотностью. Оказалось, что конъюгат конканавалина А с ферритином специфически связывается на наружной, но не на внутренней поверхности мембраны эритроцитов. Такая же асимметрия наблюдалась и в отношении связывания многих других лекти-нов мембранами различных клеток. Таким образом, наружная локализация углеводных единиц анионного канала и гликофорина (рис. 10.23 и 10.24) отражает общий принцип строения мембран, а именно: остатки Сахаров локализованы на наружной поверхности плазматической мембраны. Во всех исследованных к настоящему времени клетках млекопитающих остатки сахаров
-маннозы, тогда как агглютинин из зародыша пшеницы присоединяется к концевым остаткам N-ацетилглюкозамина. Эти лектины хорошо видны в электронном микроскопе, когда они соединены с ферритином - белком, содержащим гидроксид железа и потому обладающим высокой электронной плотностью. Оказалось, что конъюгат конканавалина А с ферритином специфически связывается на наружной, но не на внутренней поверхности мембраны эритроцитов. Такая же асимметрия наблюдалась и в отношении связывания многих других лекти-нов мембранами различных клеток. Таким образом, наружная локализация углеводных единиц анионного канала и гликофорина (рис. 10.23 и 10.24) отражает общий принцип строения мембран, а именно: остатки Сахаров локализованы на наружной поверхности плазматической мембраны. Во всех исследованных к настоящему времени клетках млекопитающих остатки сахаров
в плазматических мембранах локализованы только на наружной стороне мембраны (рис. 10.25).
Возможно что углеводные группы служат для ориентирования гликопротеинов в мембране. Обладая ярко выраженными гидрофильными свойствми, остатки Сахаров в гликопротеинах или гликолипидах должны располагаться на поверхности мембраны, а не в ее углеводородной сердцевине. Энергетическая цена встраивания олигосахаридной цепи в углеводородное окружение внутри мембраны очень высока. Стало быть, существует барьер, препятствующий свободному вращению гликопротеина от одной стороны мембраны к другой. Углеводные компоненты мембранных гликопротеинов способствуют поддержанию асимметрии биологических мембран.
Углеводы на поверхности клетки могут играть также важную роль в межклеточном узнавании. От узнавания клетками друг друга зависят такие, например, процессы, как формирование тканей путем взаимодействия различных клеток или распознавание чужеродных клеток иммунной системой высших организмов.


Рис. 10.23.
Углеводы обладают потенциальной возможностью к созданию огромного структурного разнообразия. Так, набор углеводов на поверхности клеток имеет колоссальное число вариантов потому, что: 1) моносахариды могут соединяться друг с другом через любой из своих гидроксилов; 2) связь по  может быть как
может быть как  так и
так и  -конфигурации и 3) возможно интенсивное ветвление цепи. В самом деле, из четырех Сахаров может образоваться гораздо больше разных олигосахаров, чем из четырех аминокислот - разных олигопептидов.
-конфигурации и 3) возможно интенсивное ветвление цепи. В самом деле, из четырех Сахаров может образоваться гораздо больше разных олигосахаров, чем из четырех аминокислот - разных олигопептидов.