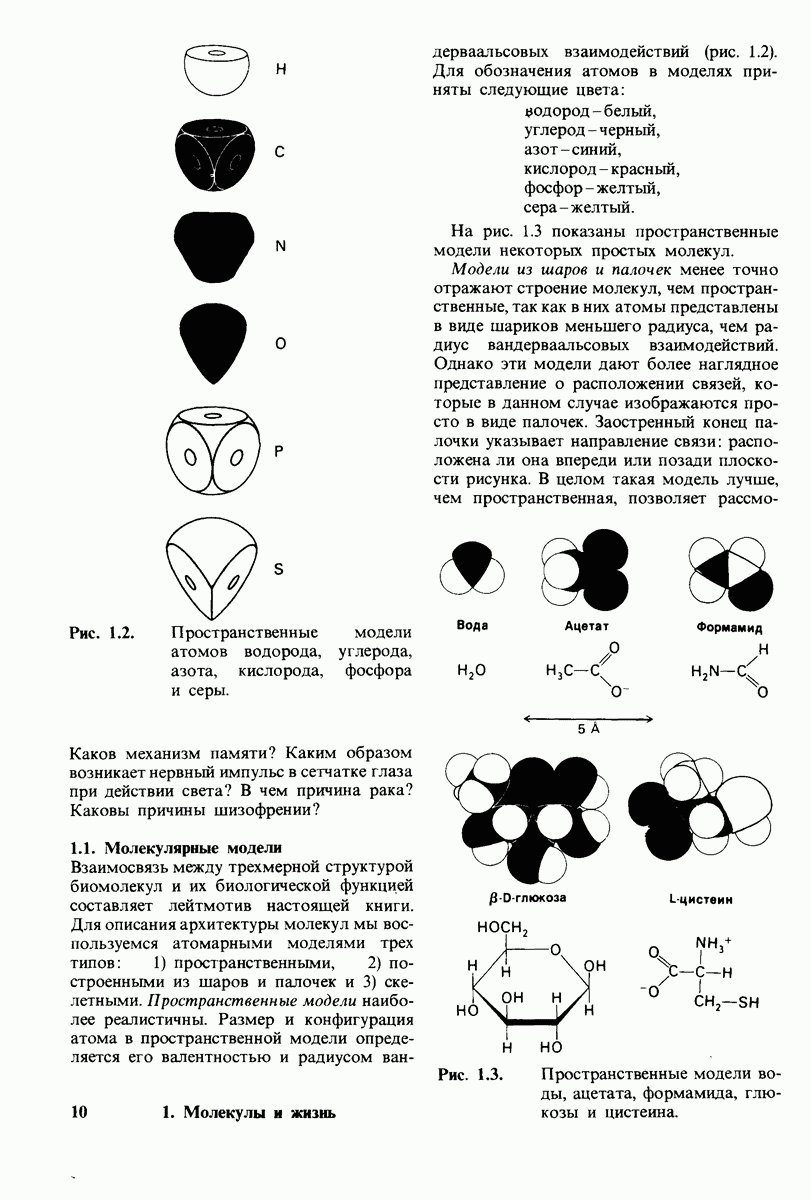
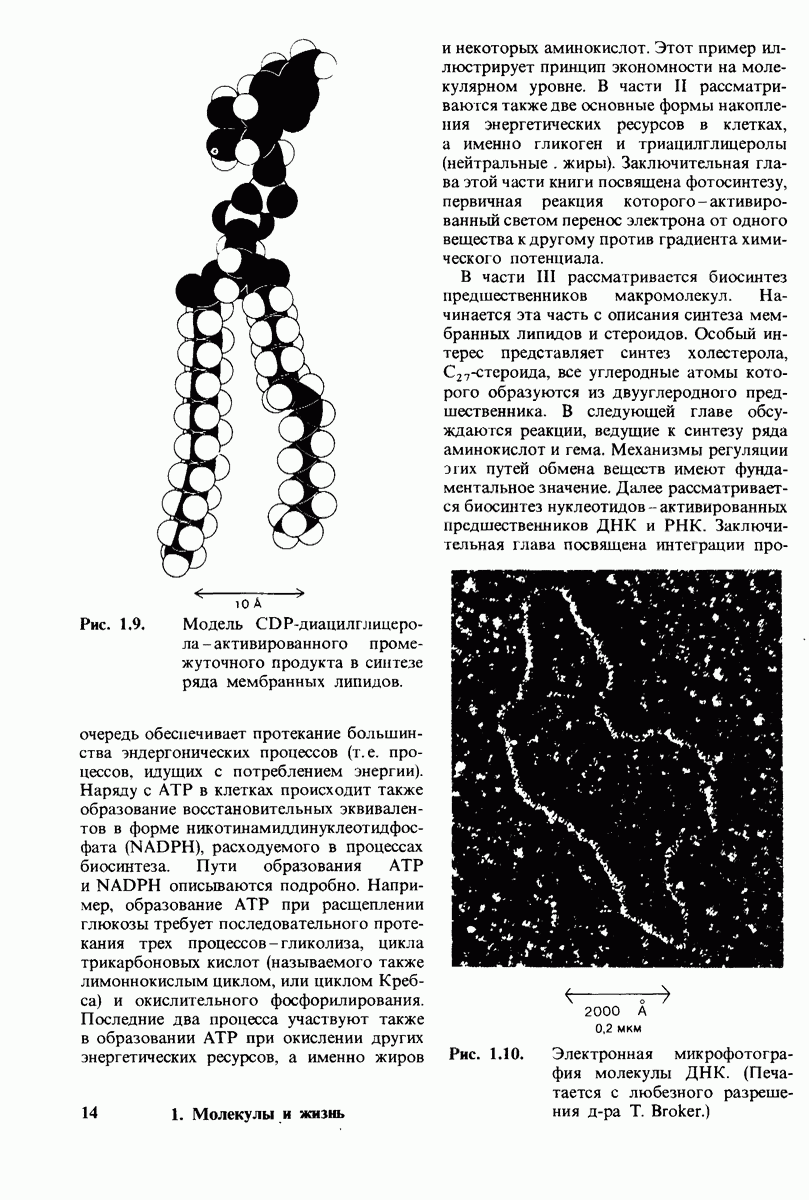
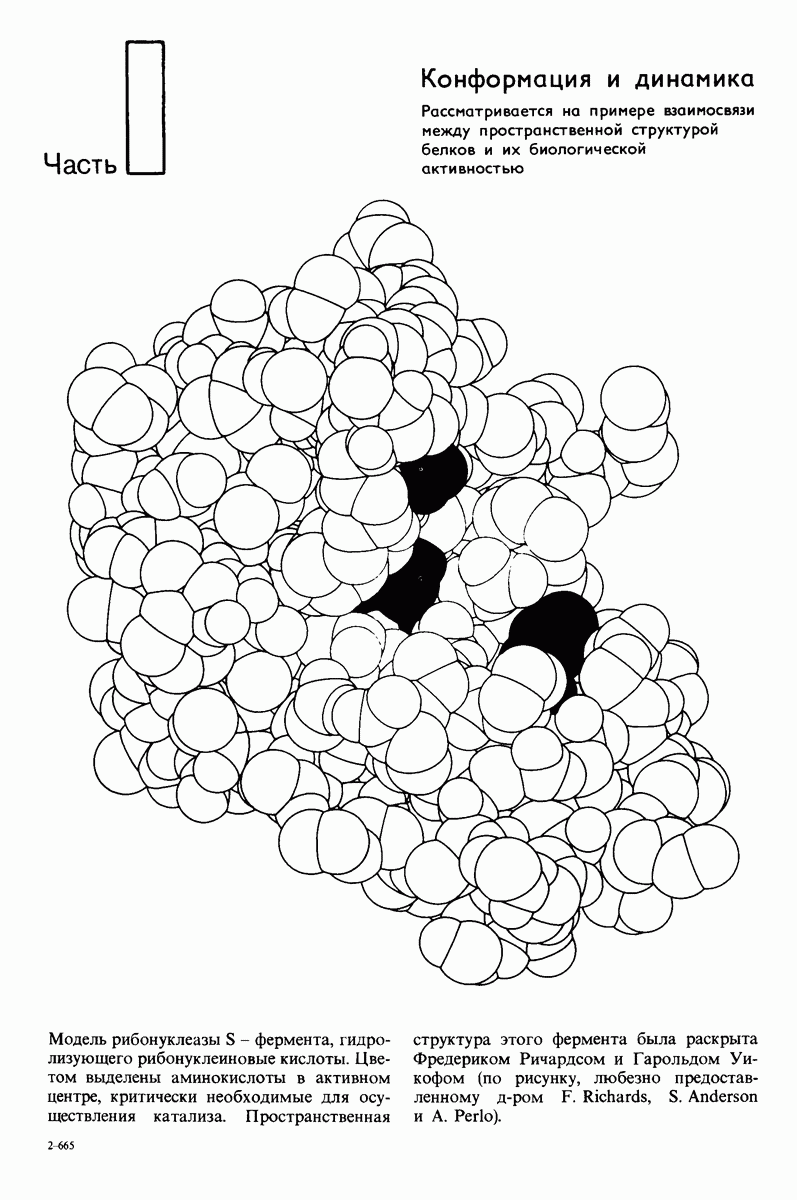
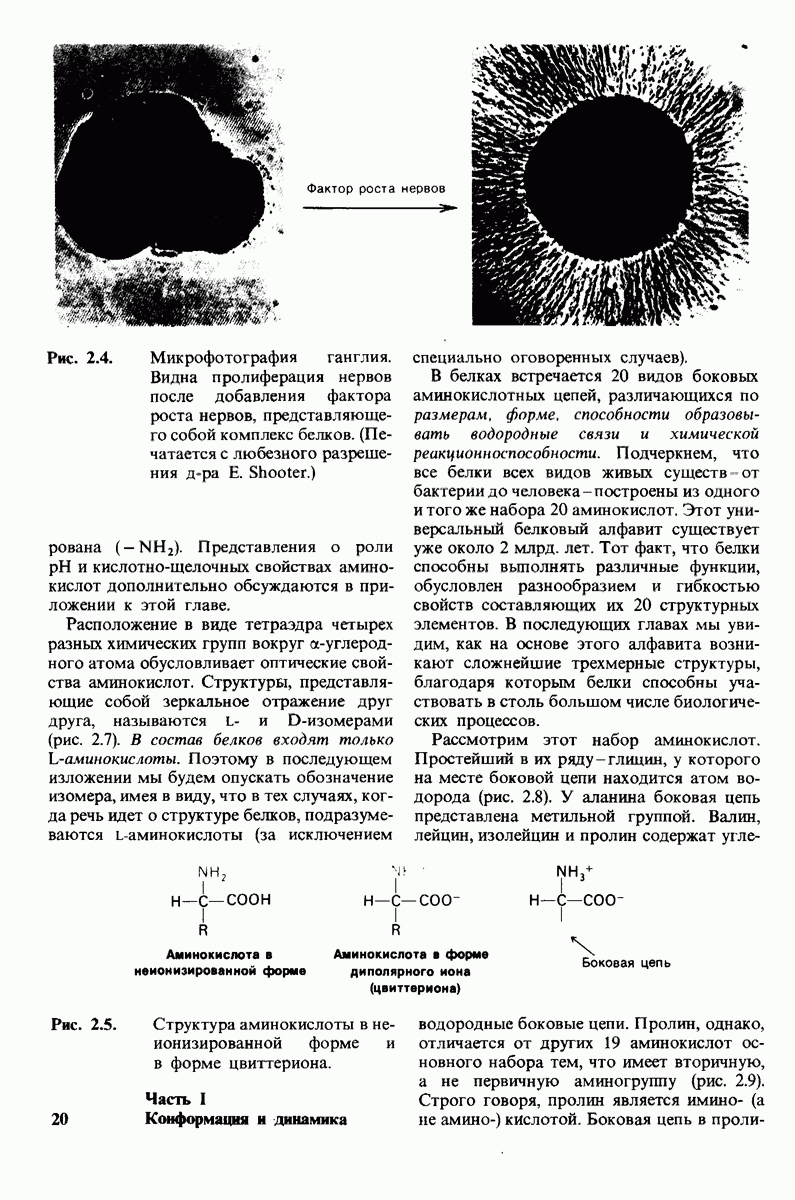
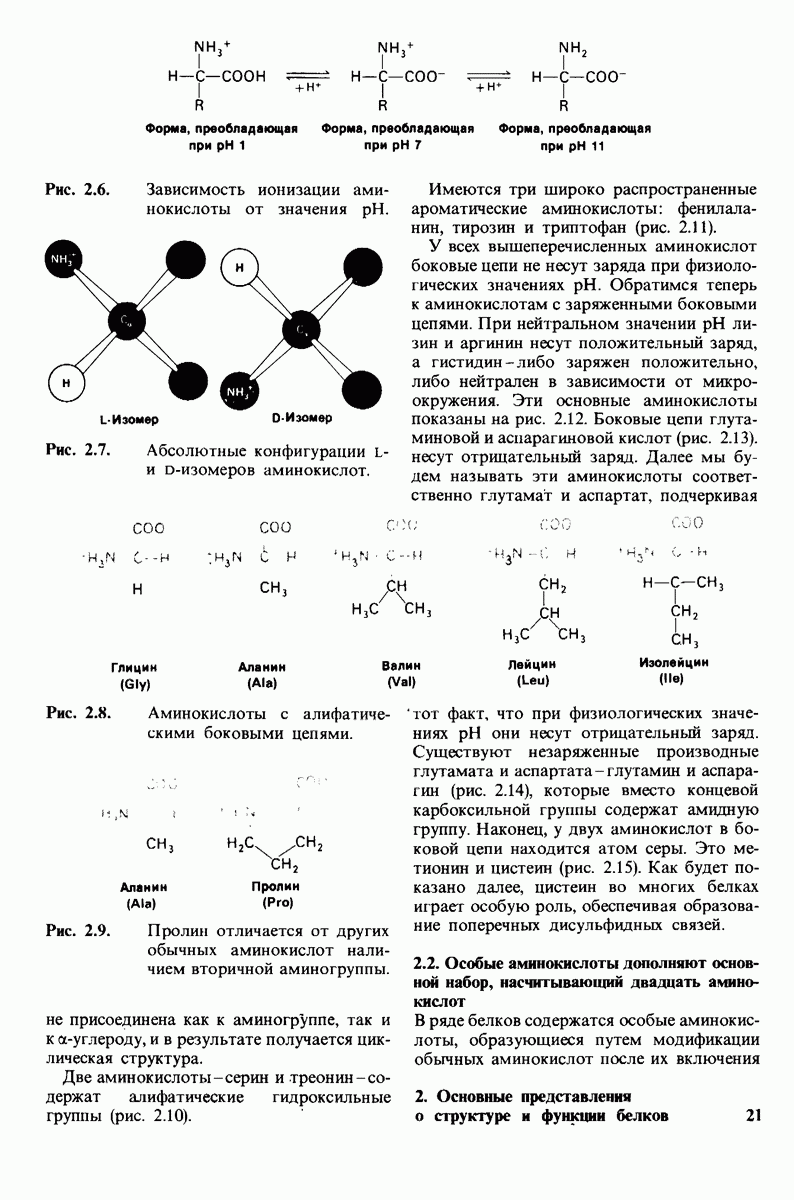
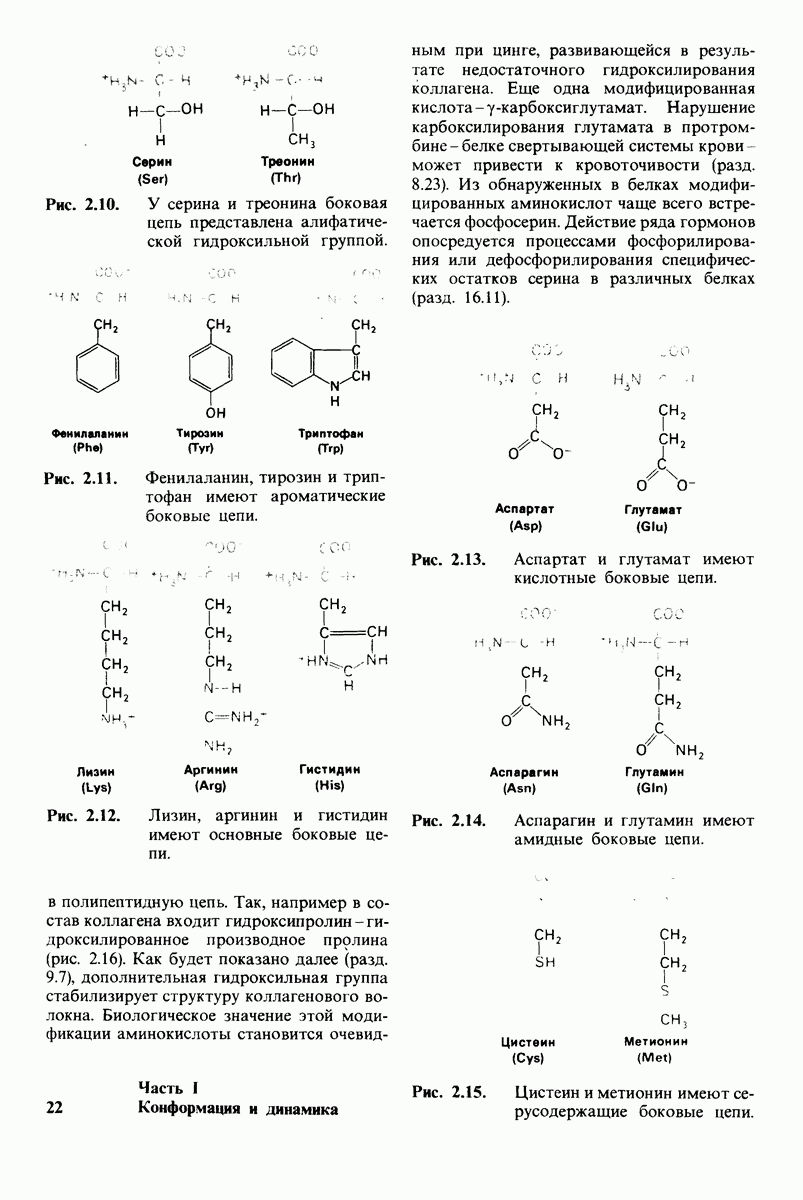
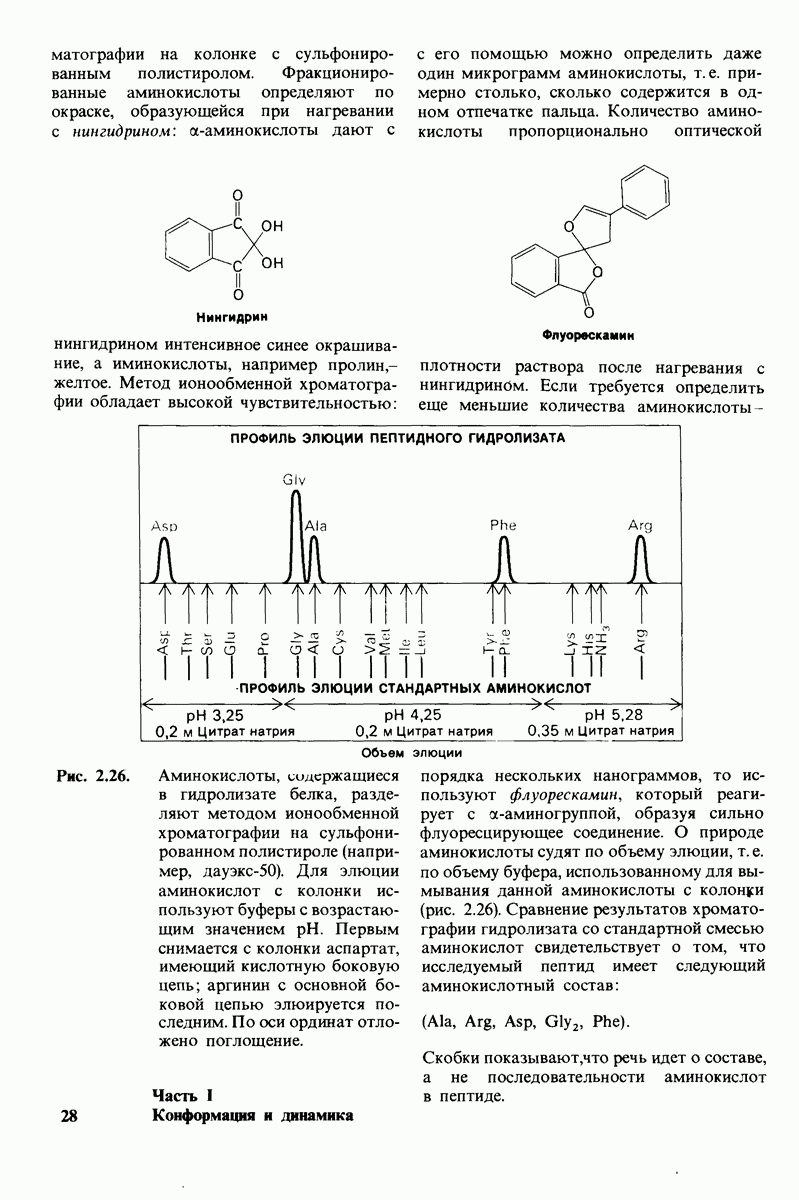
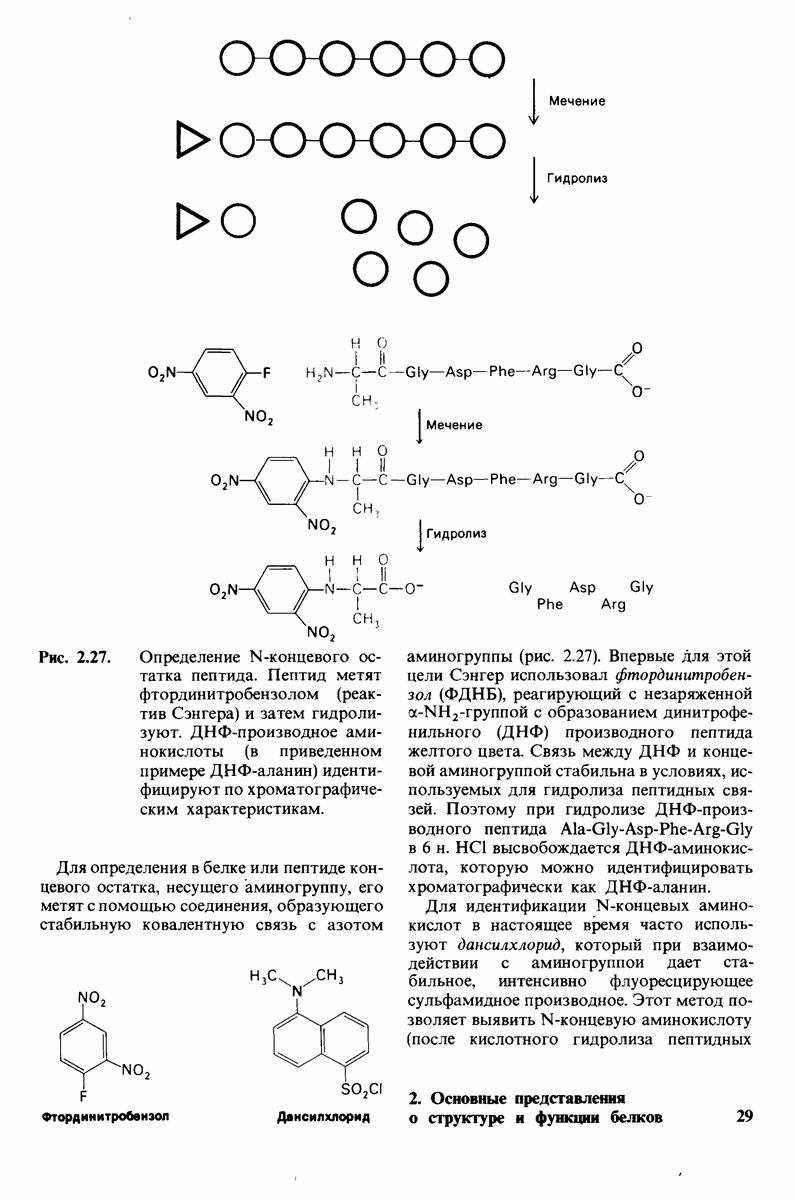
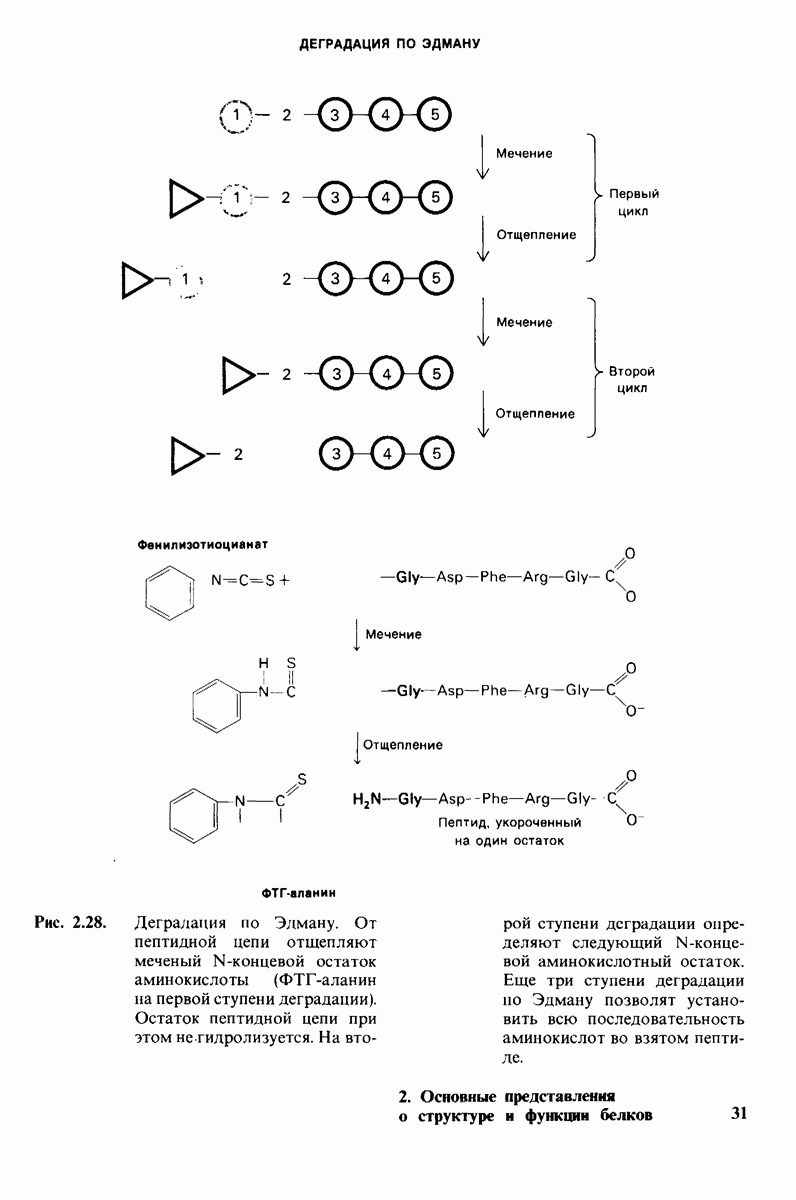
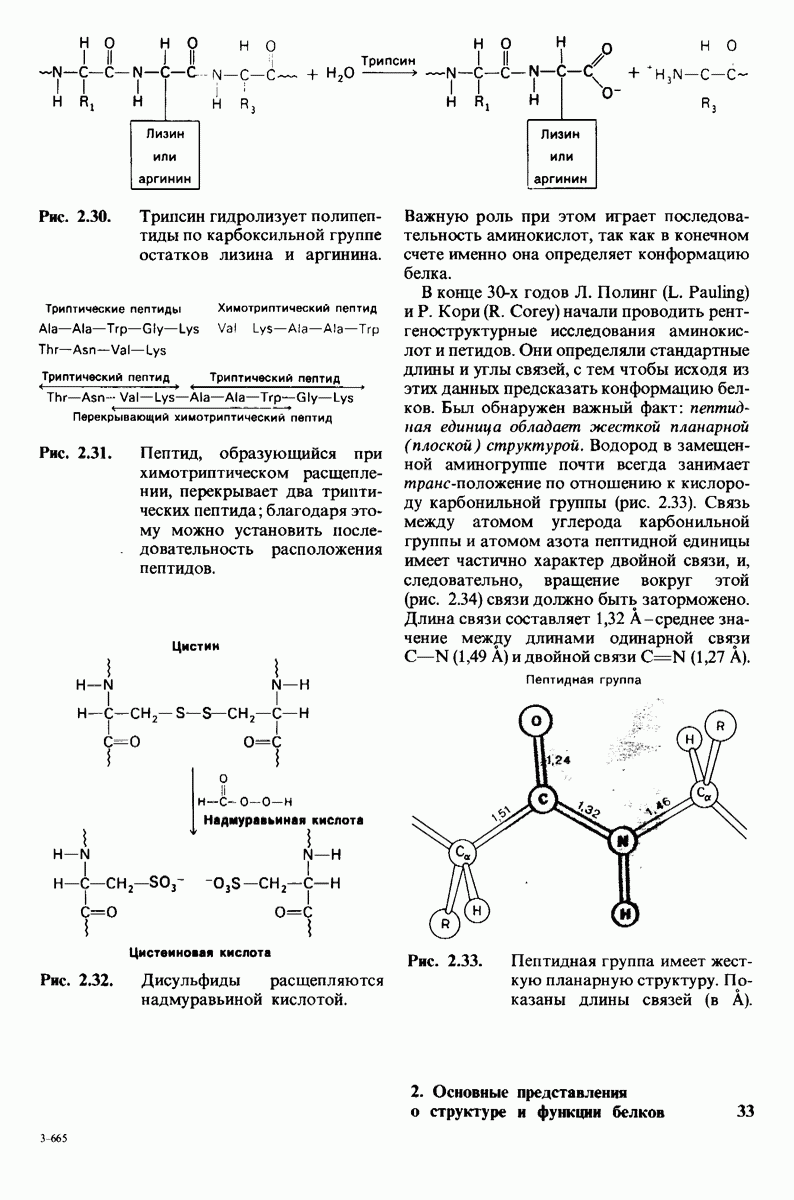
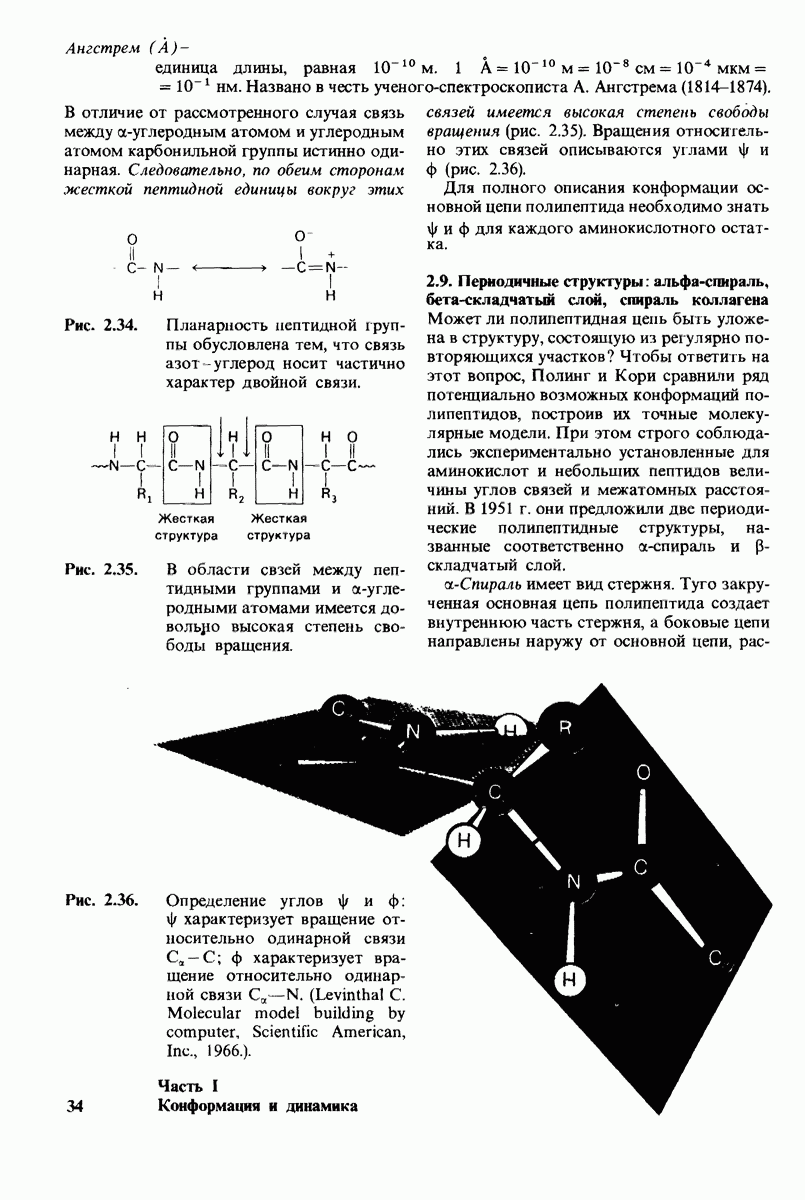
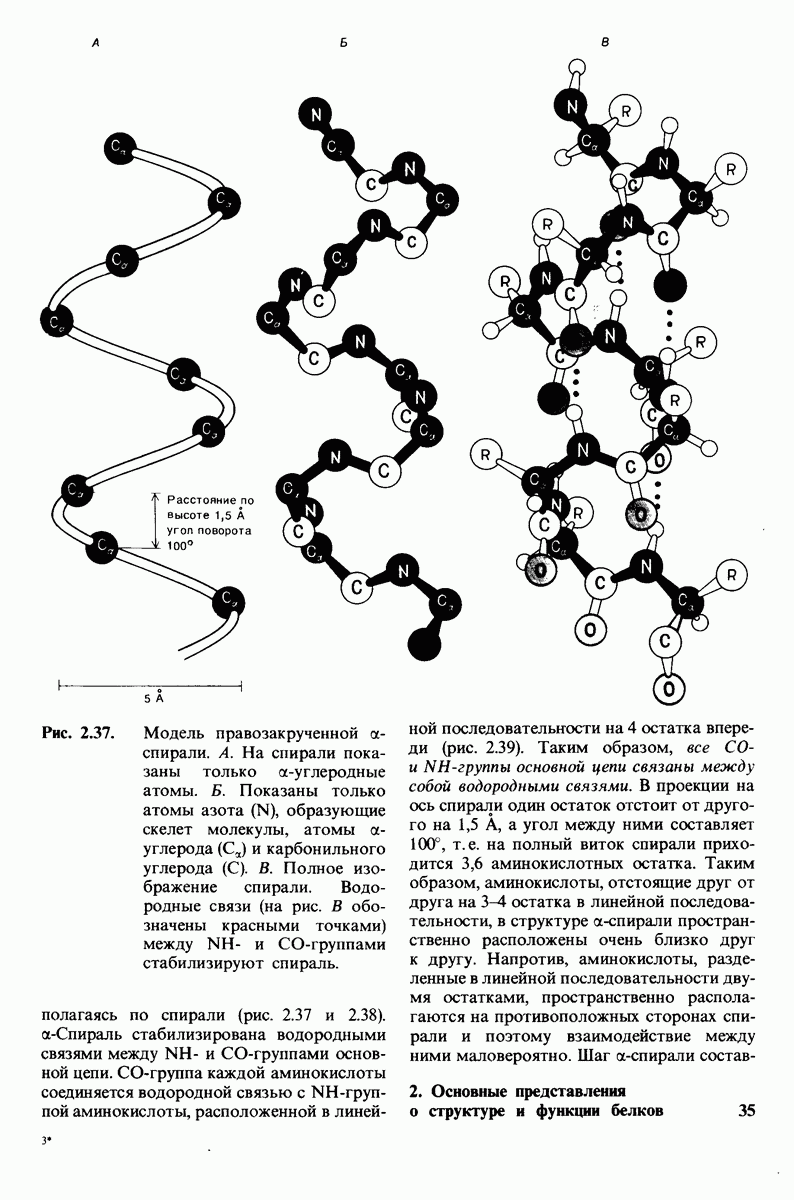
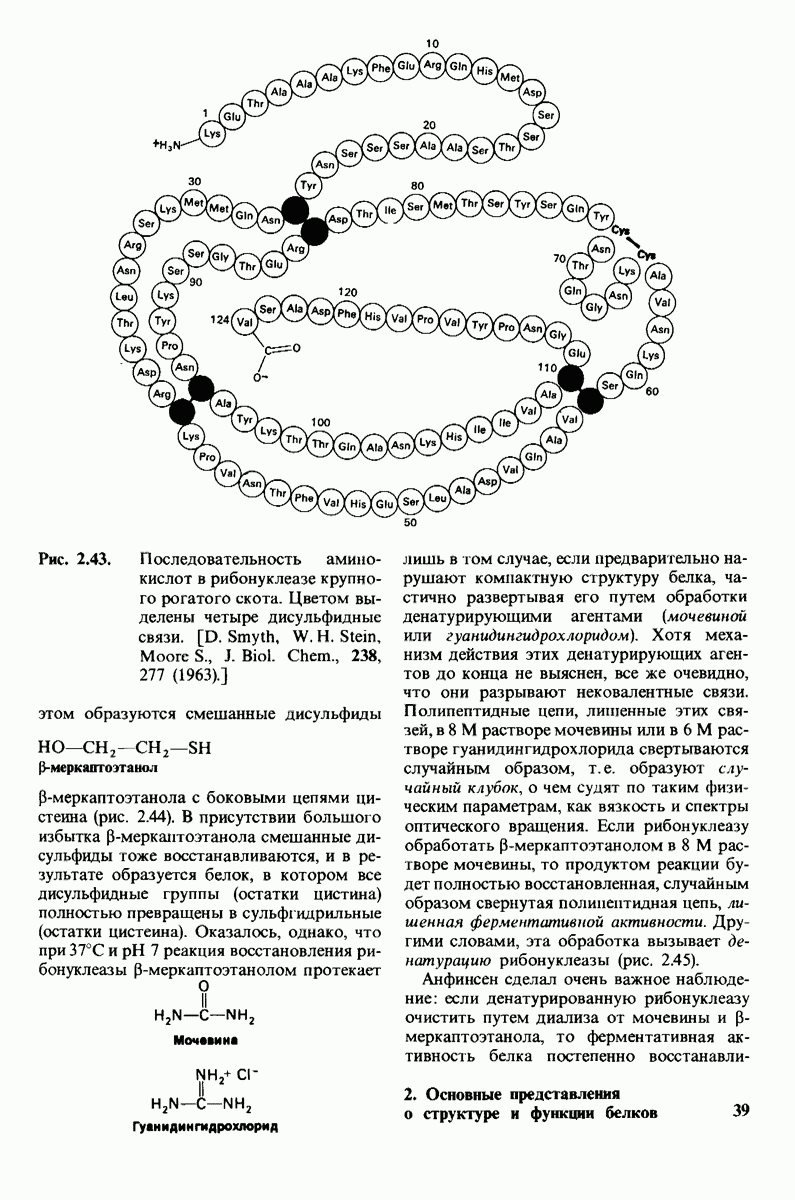
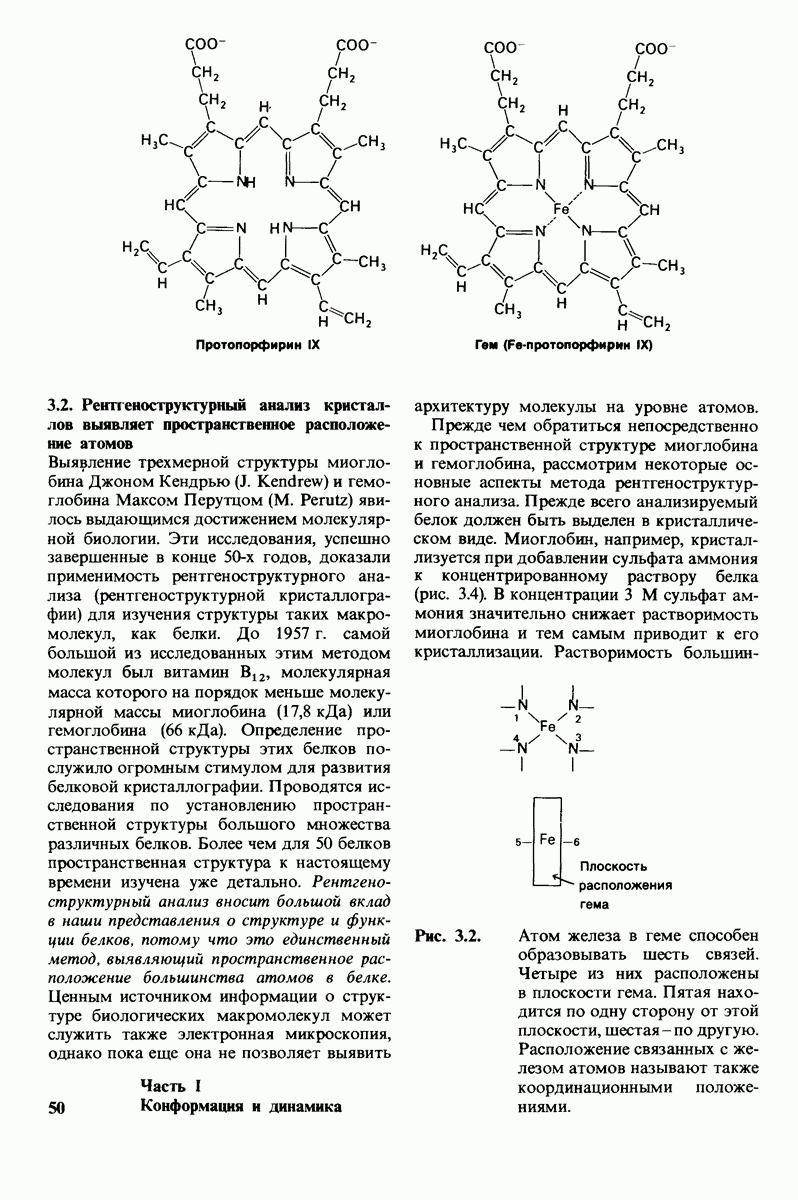
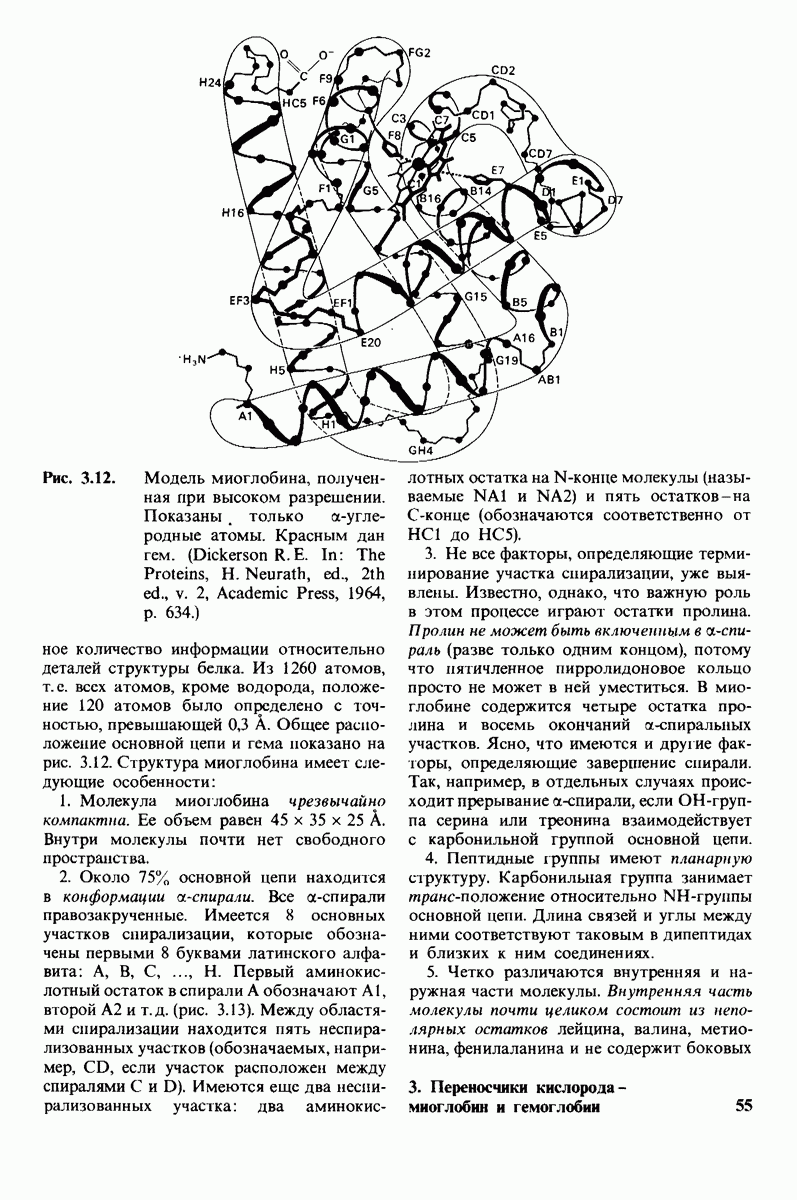
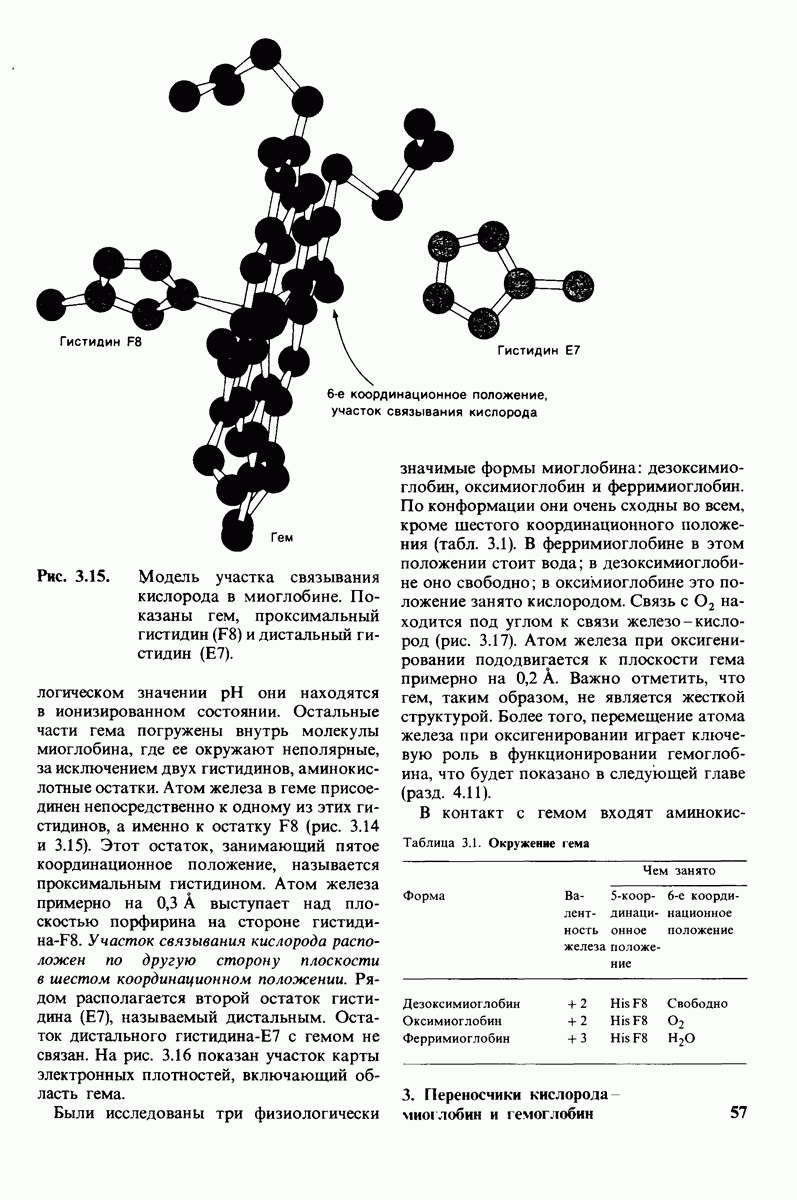
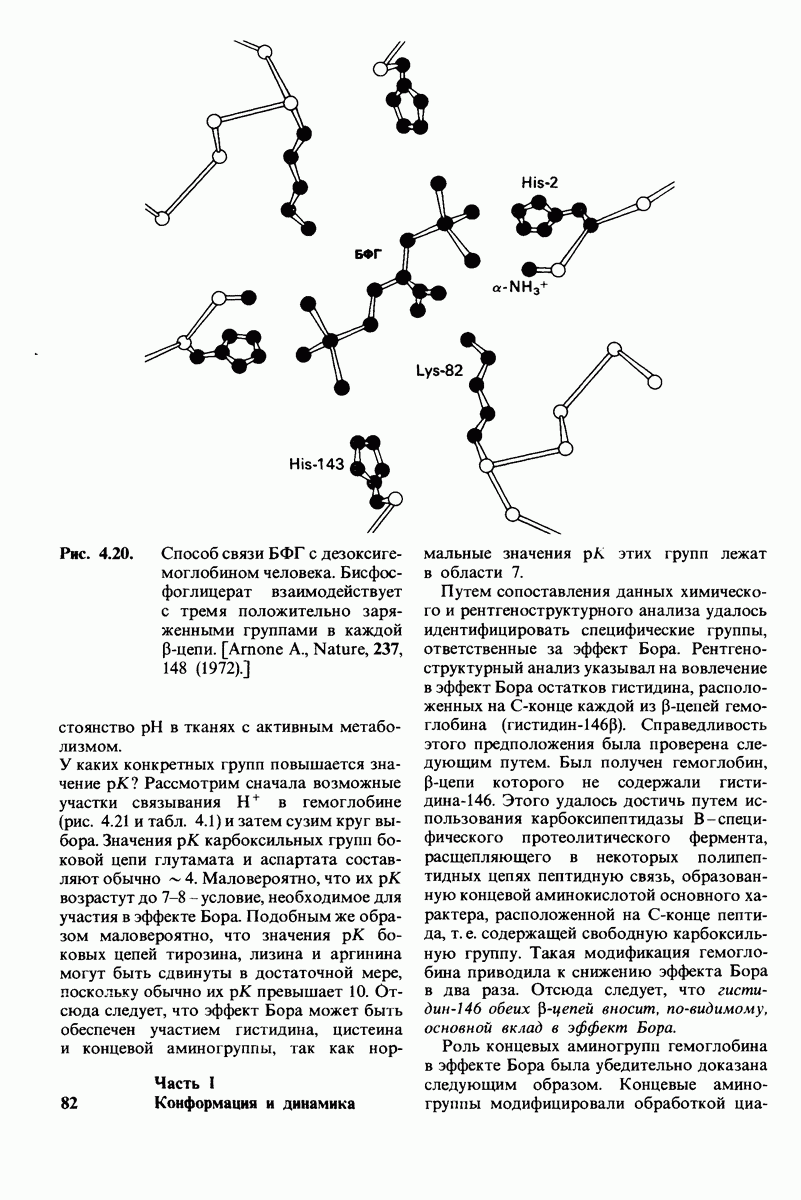
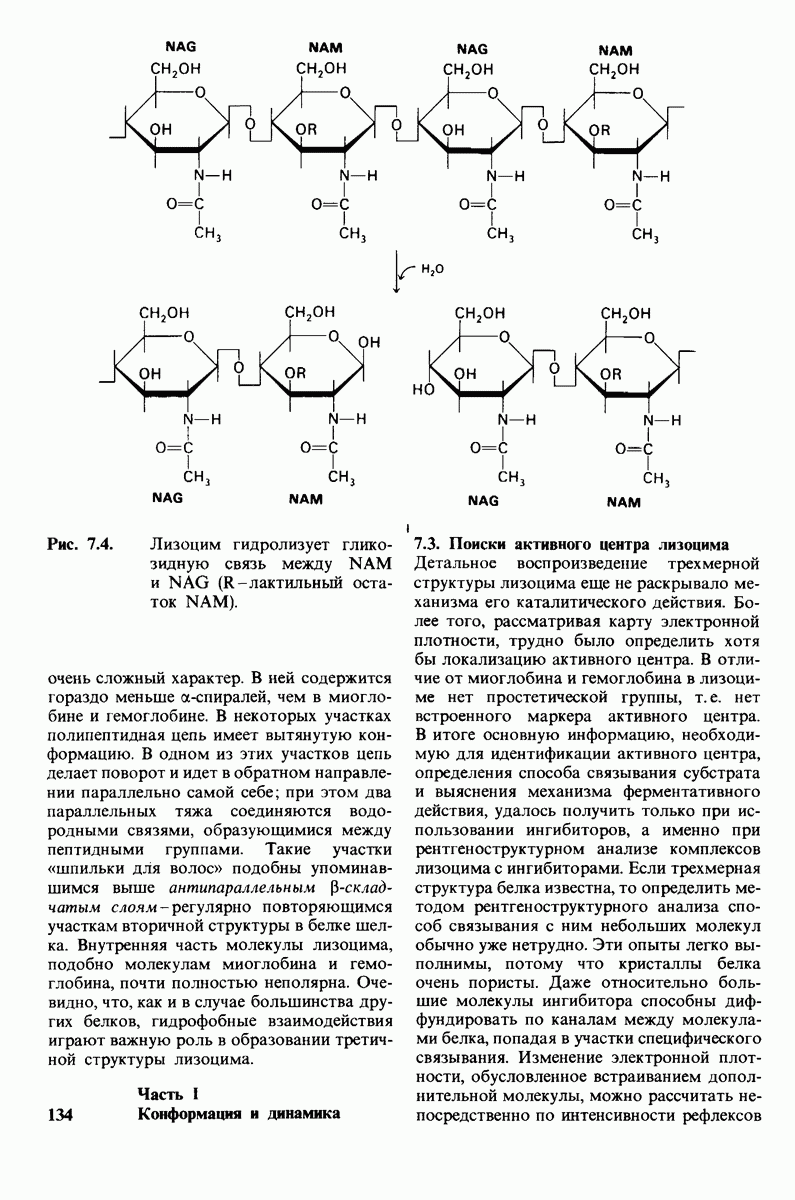
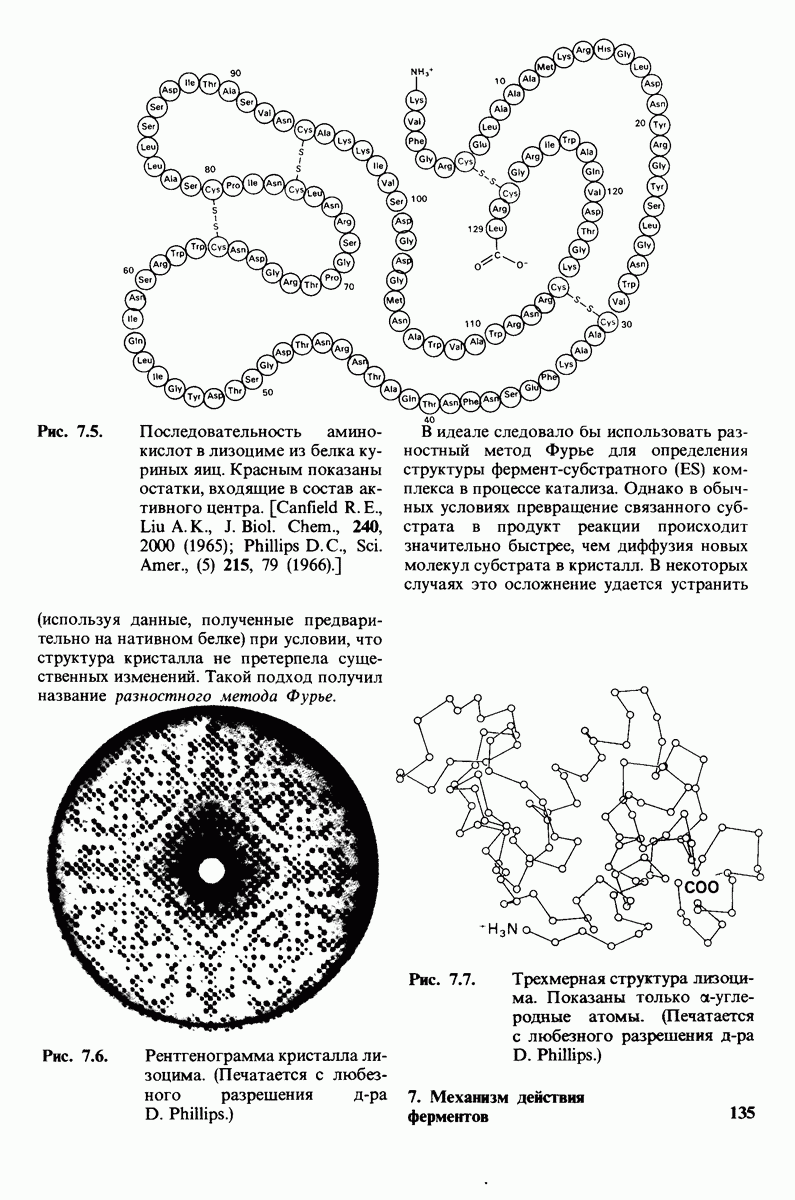
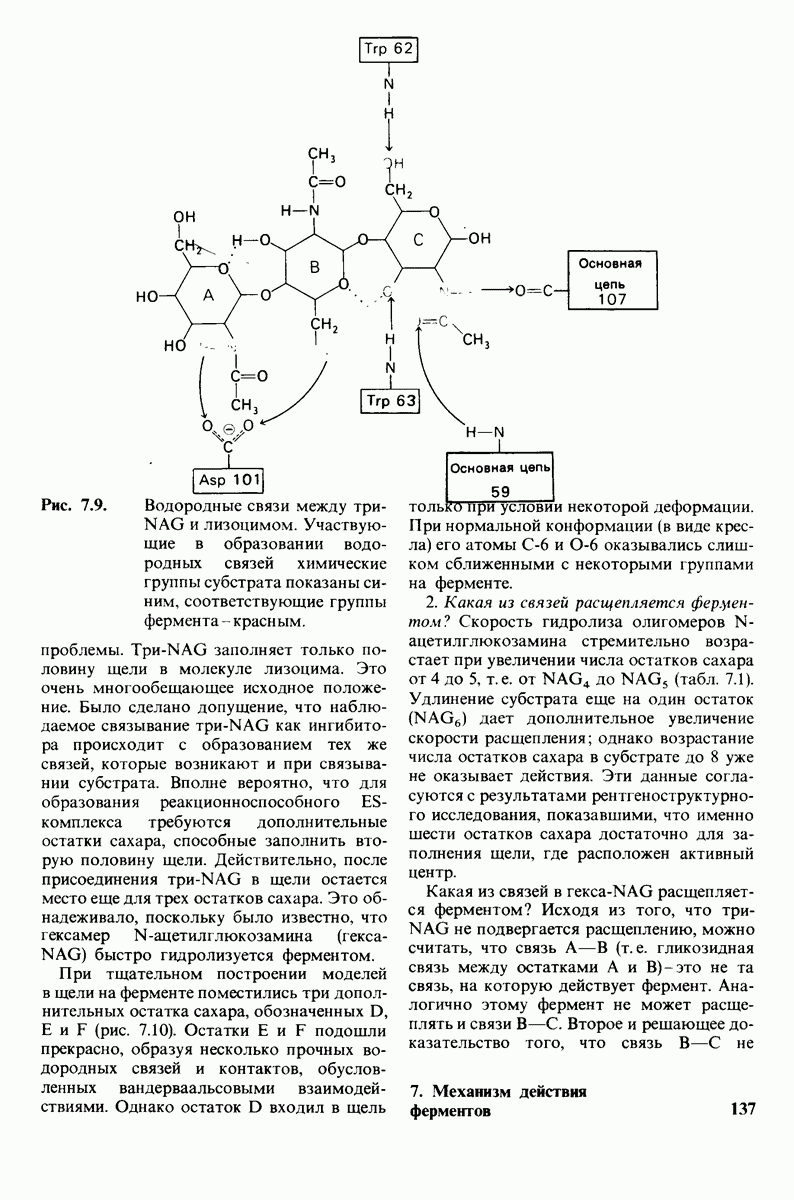
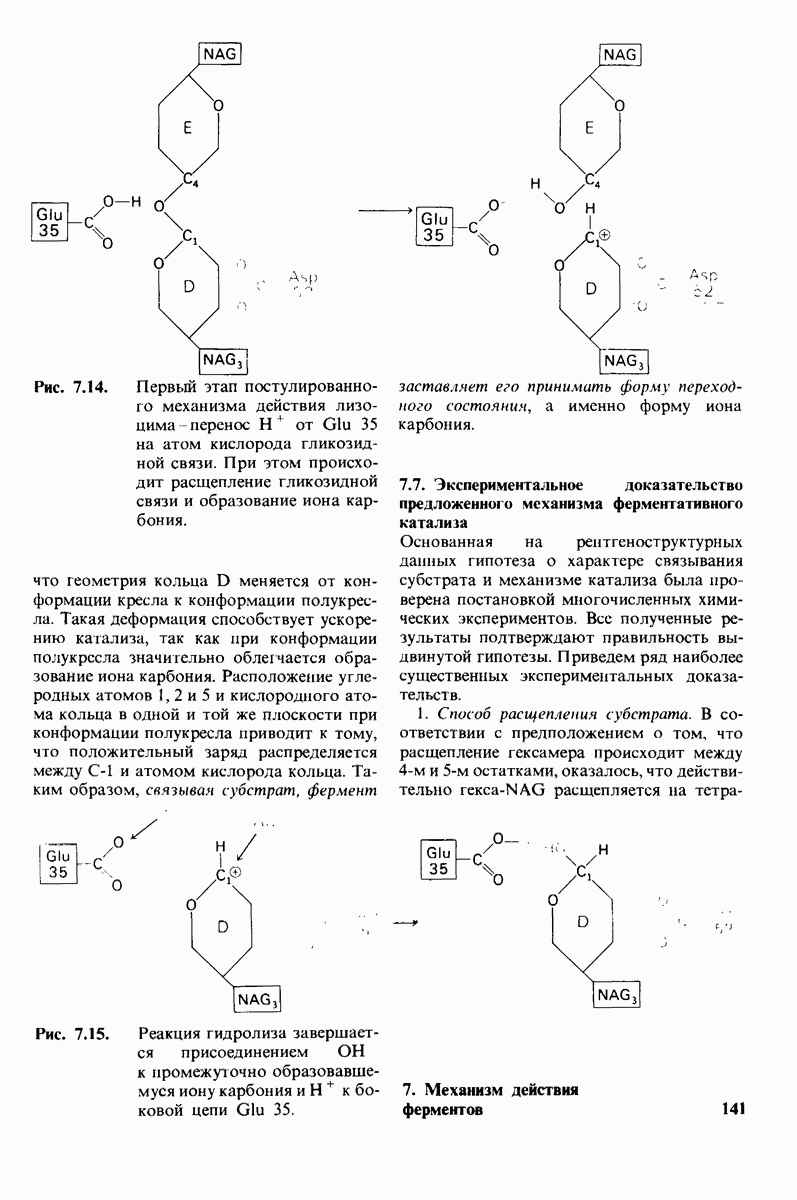
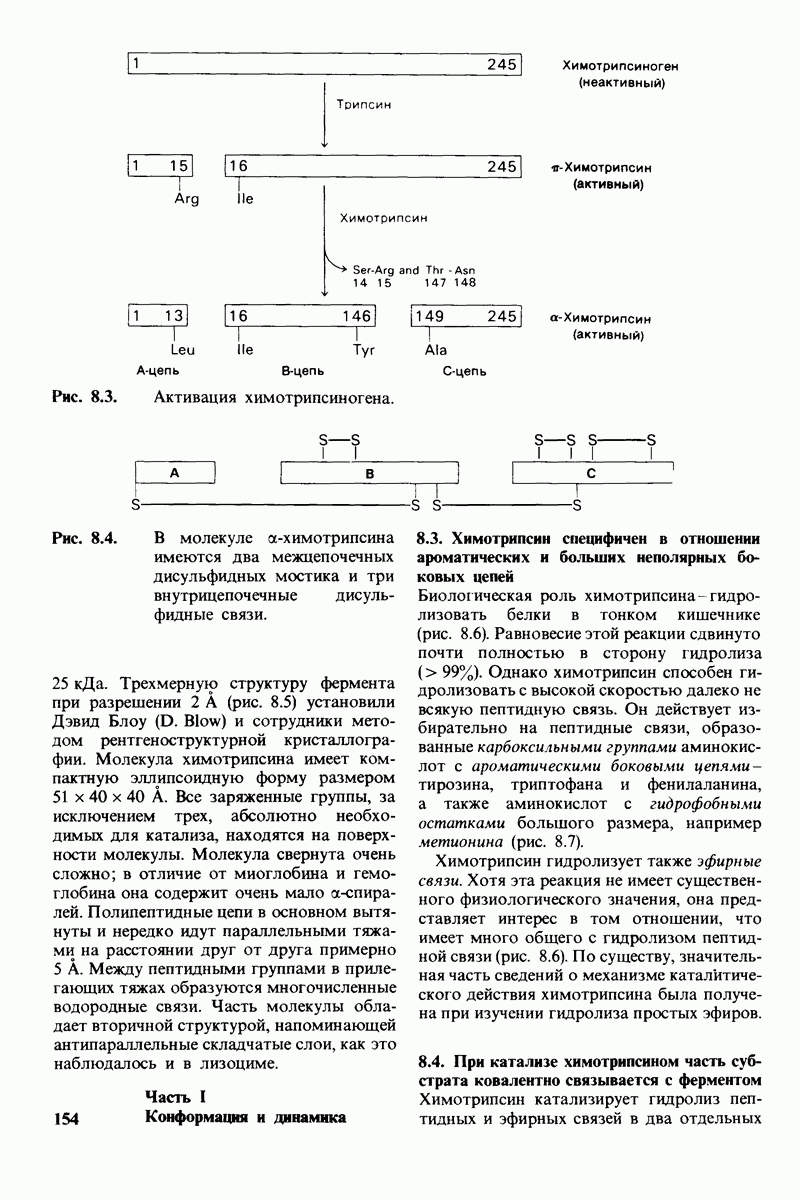
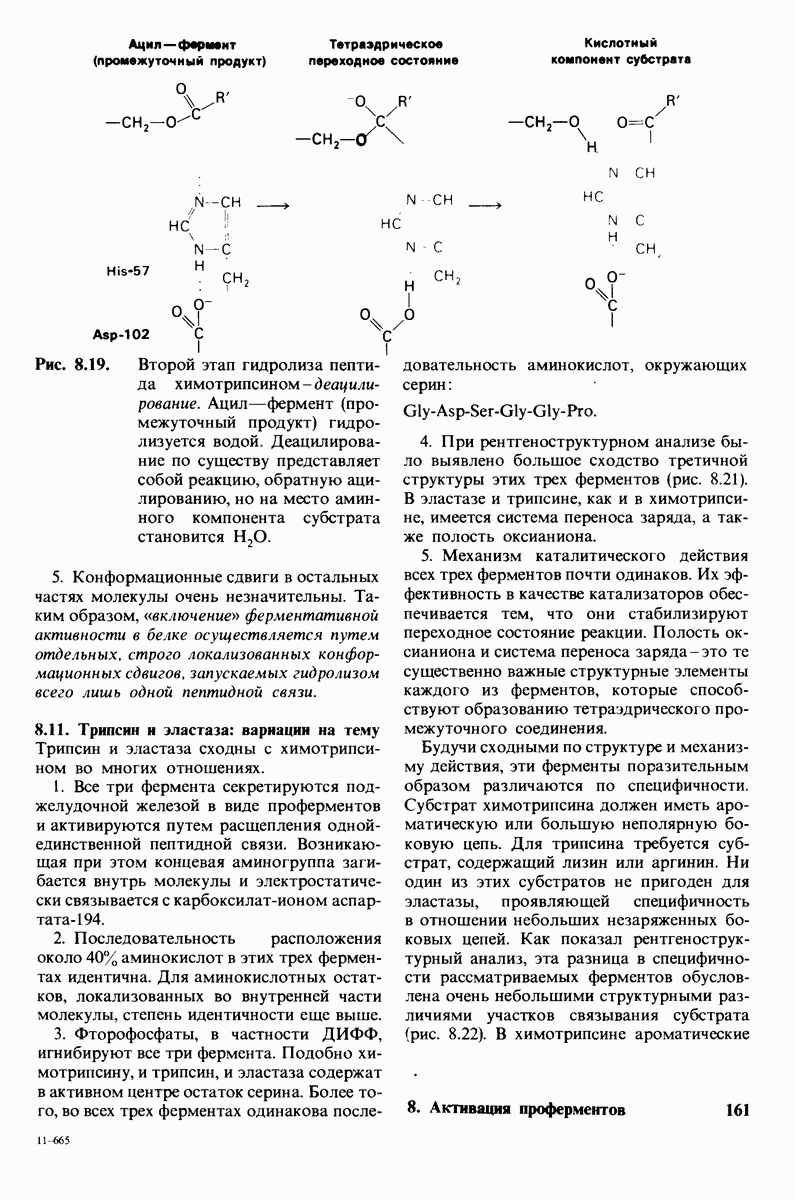
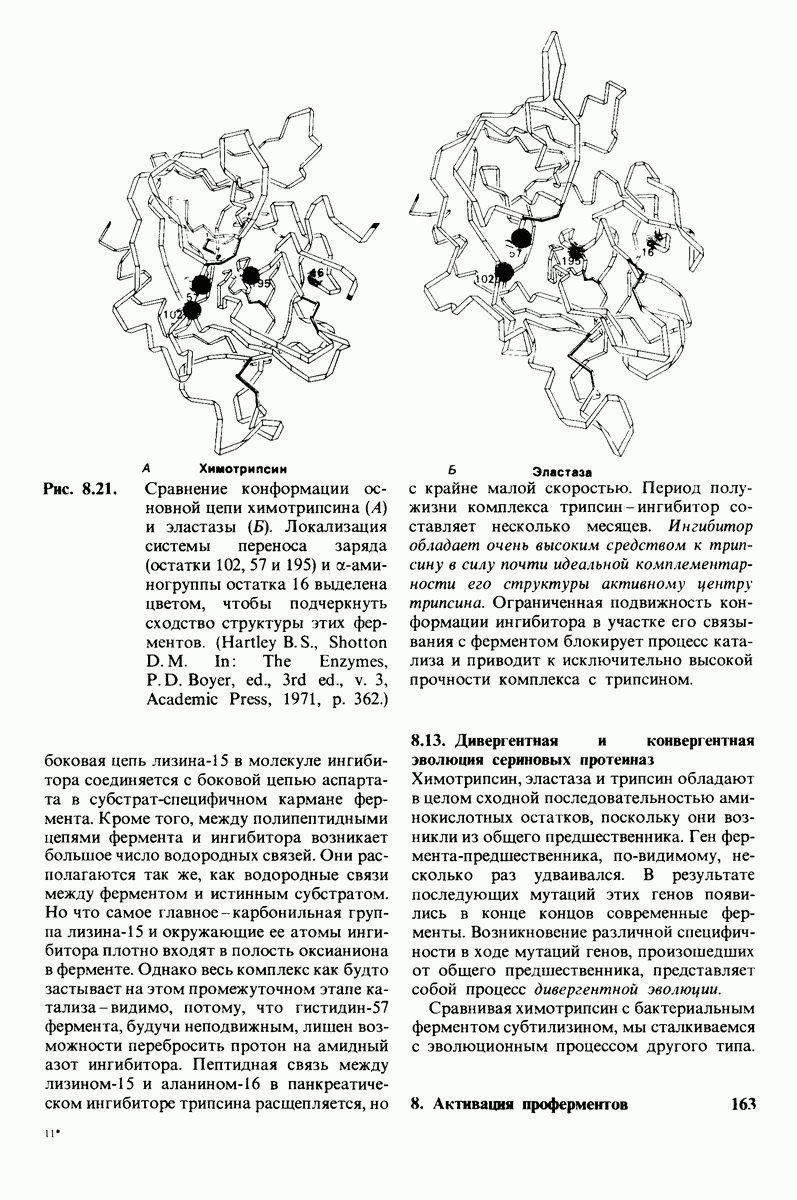
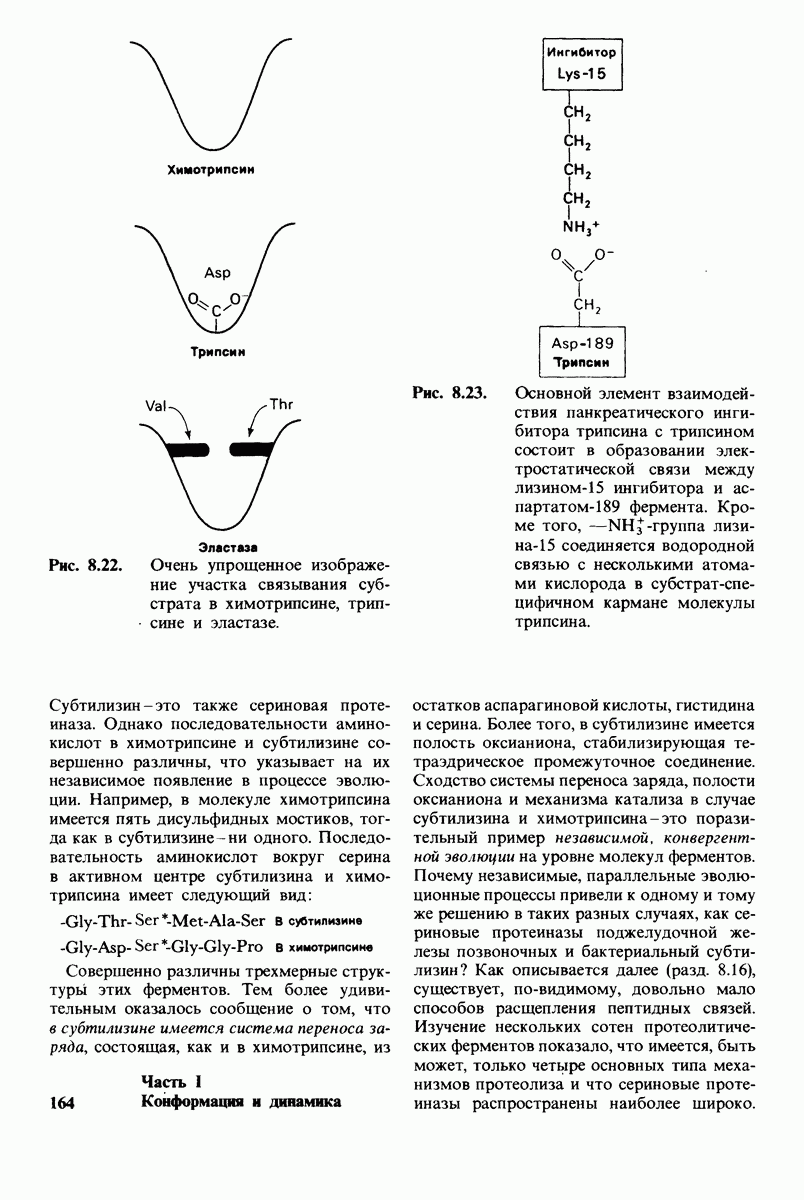
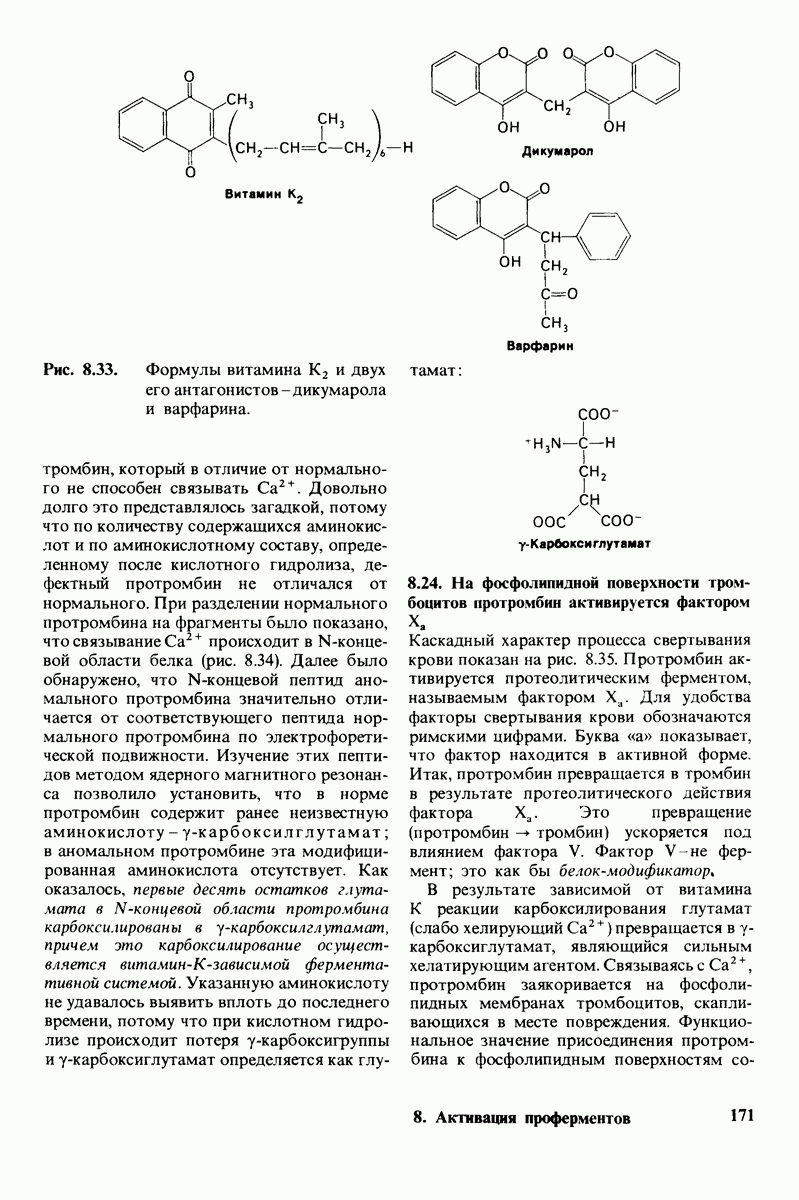
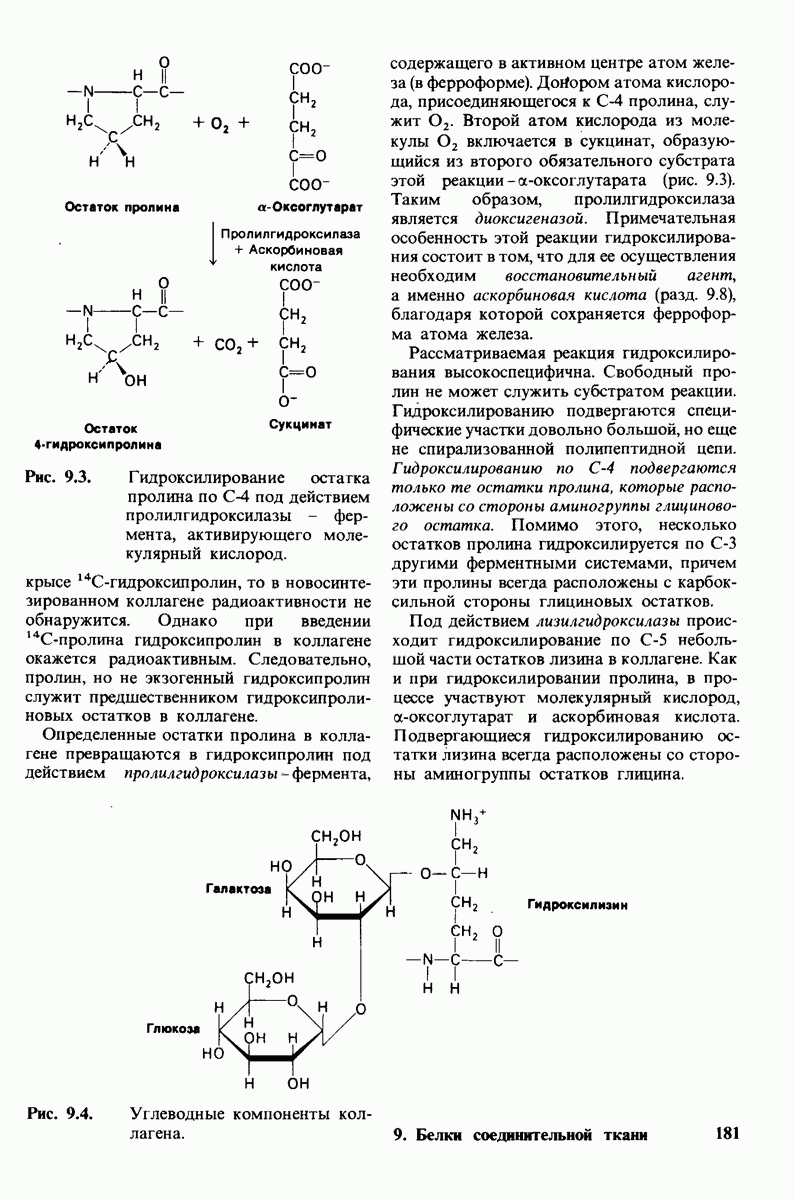
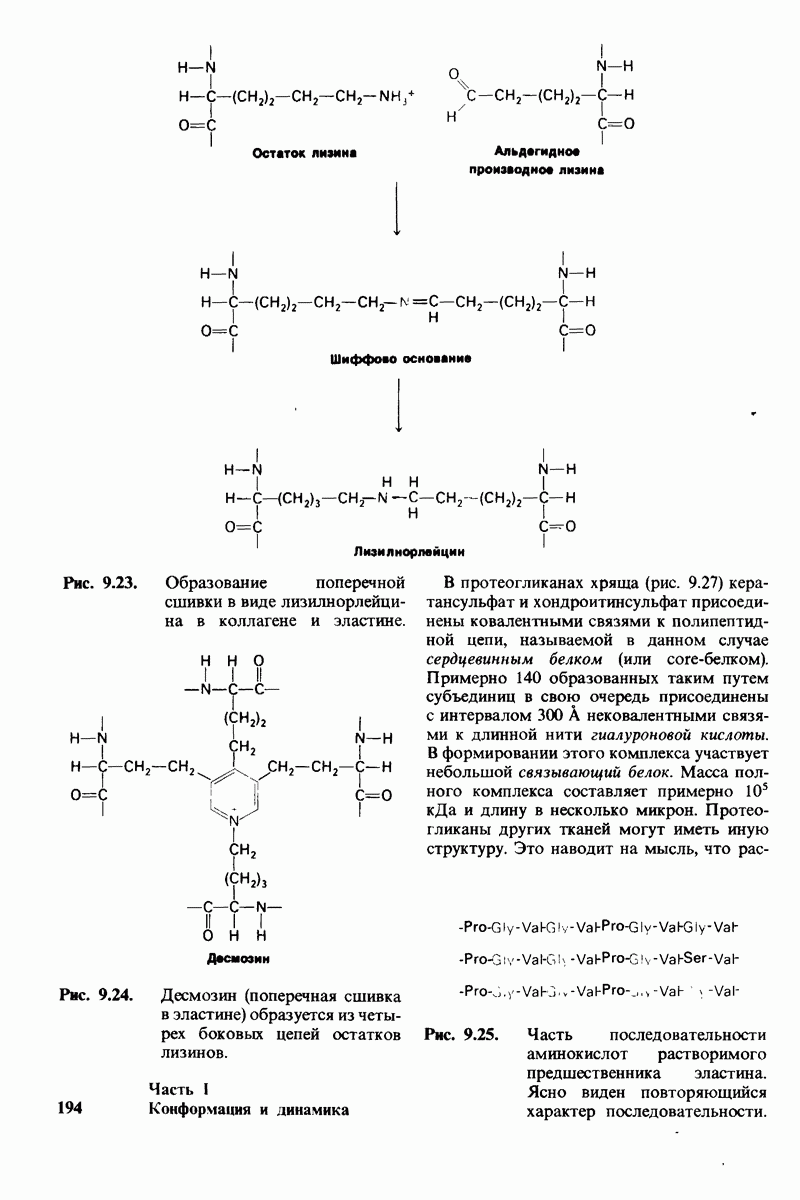
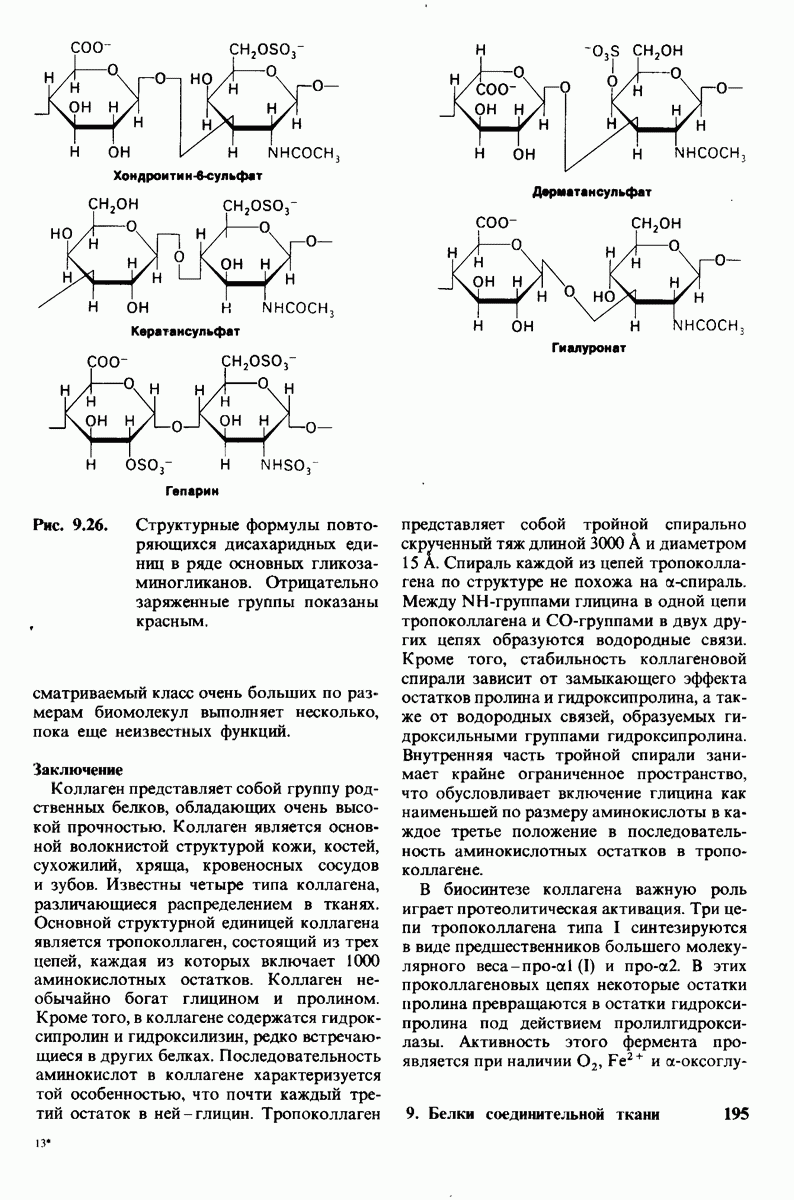
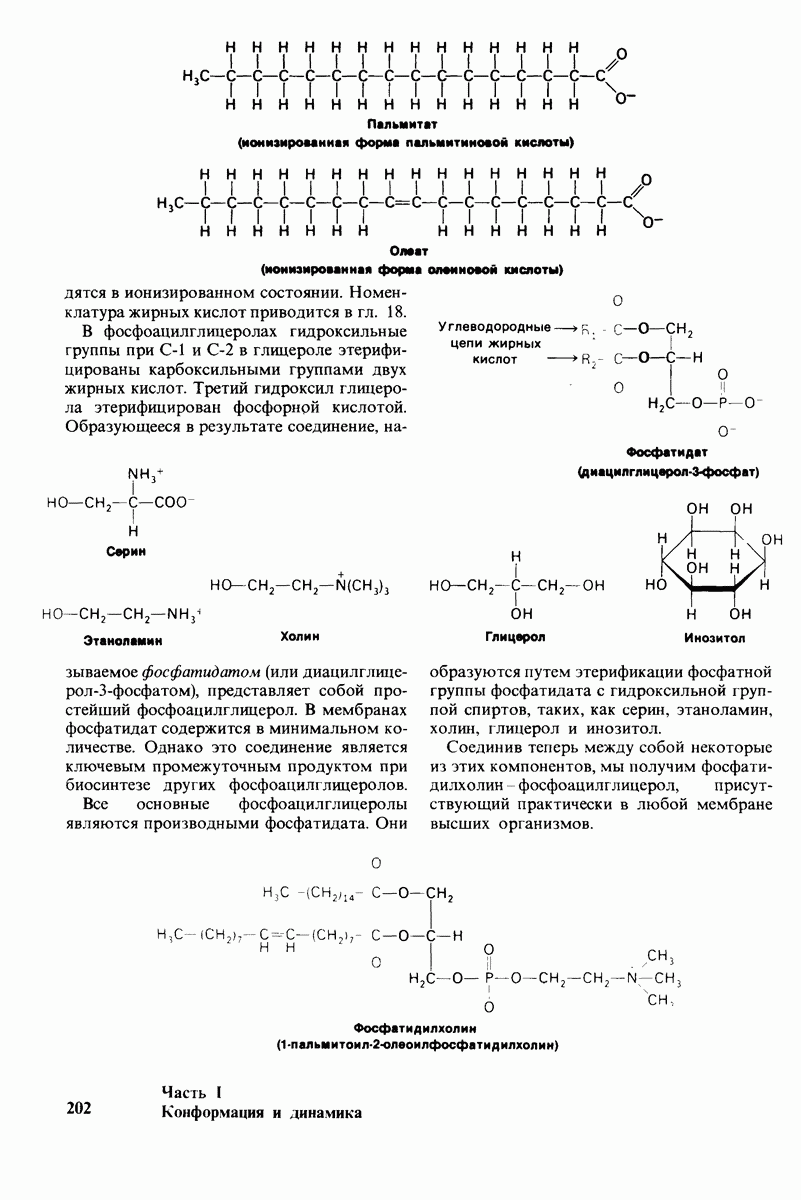
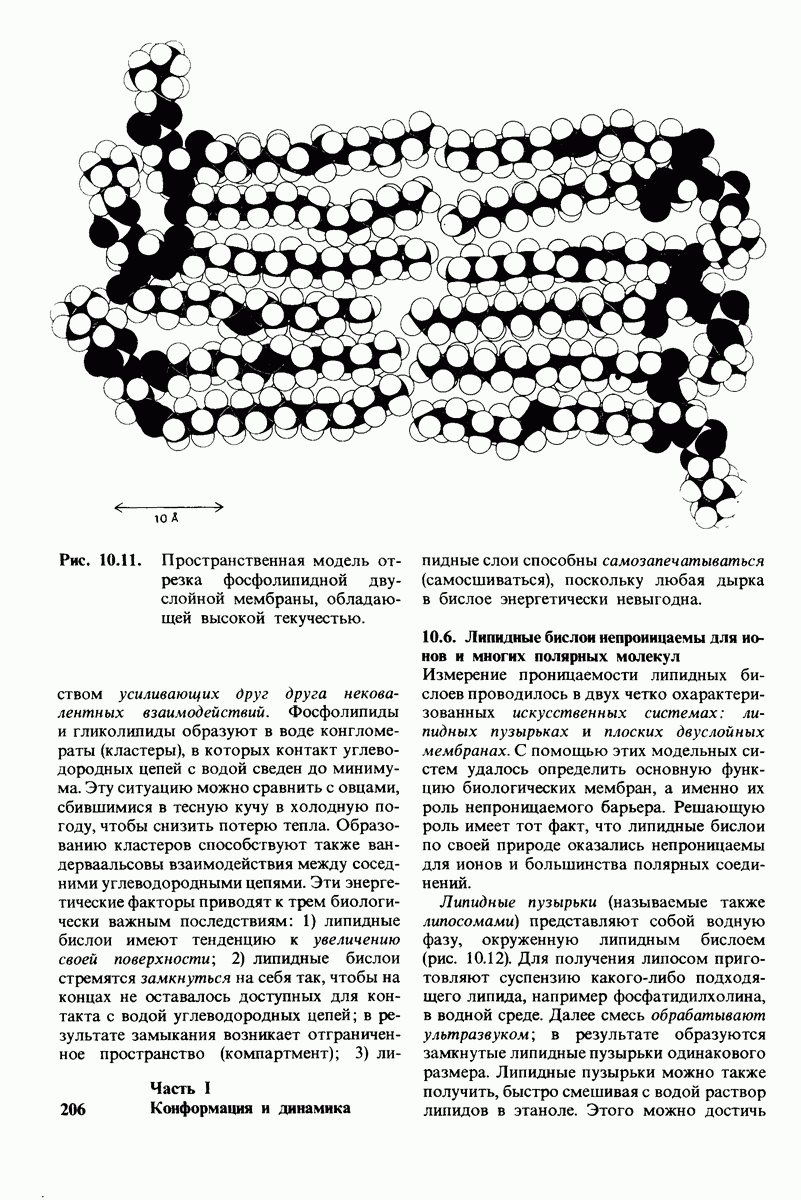
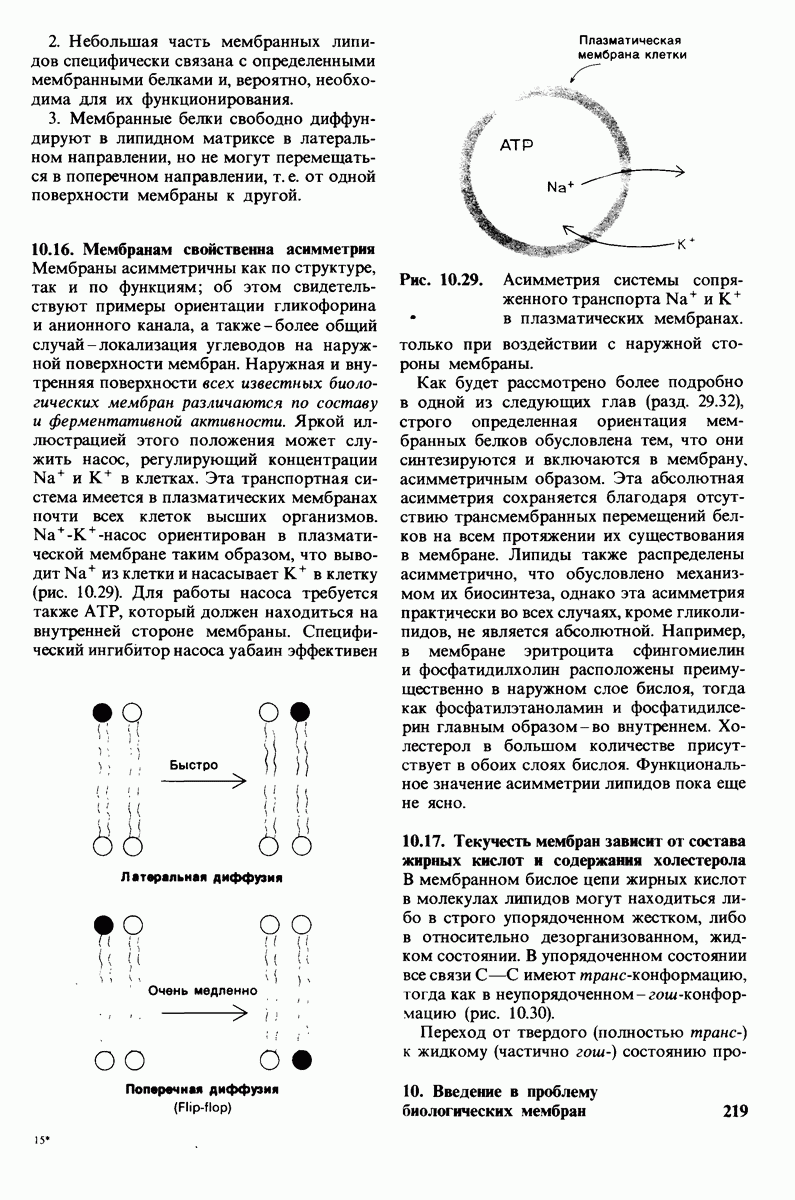
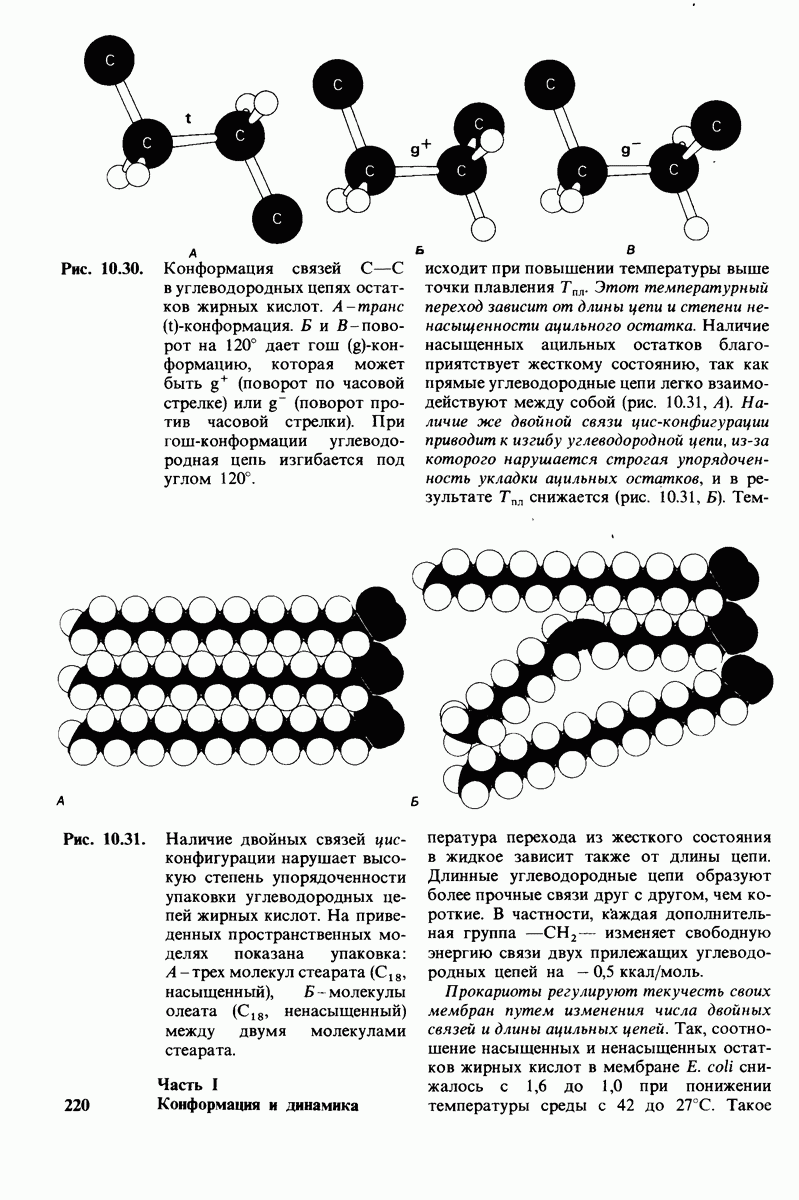
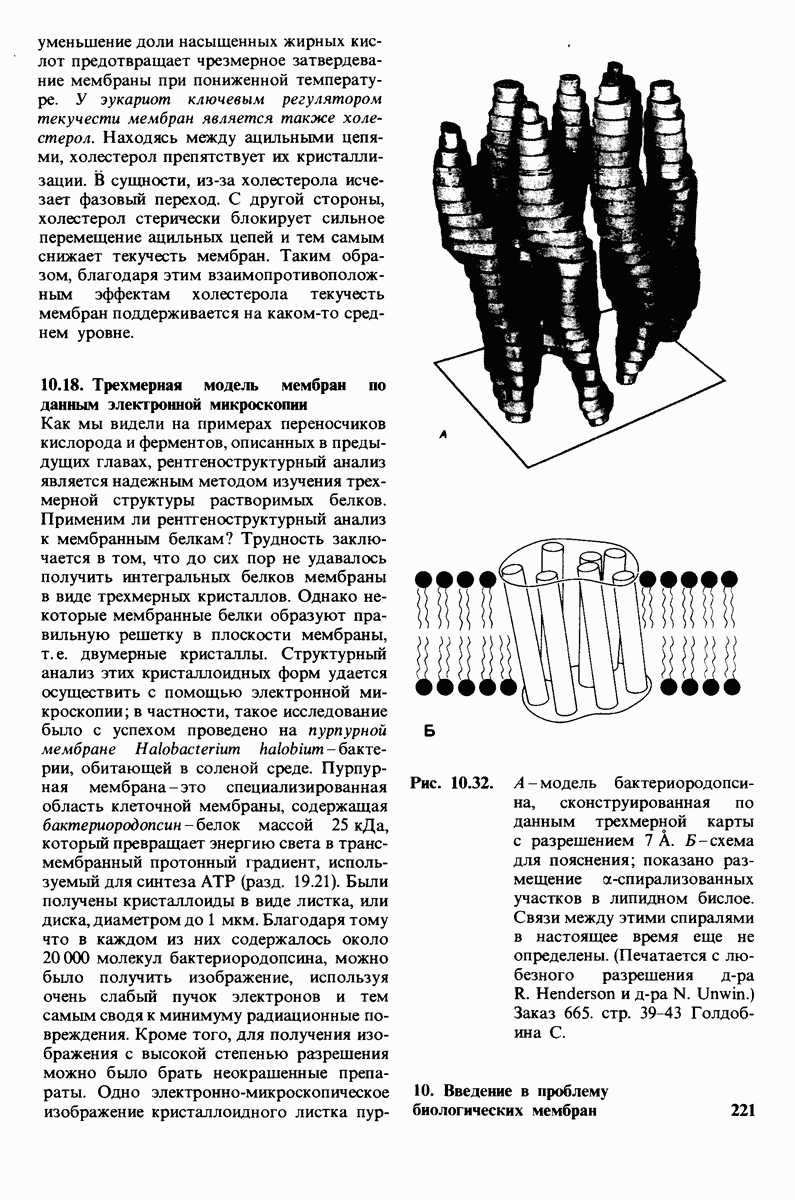
7.9. Связывание субстрата индуцирует большие структурные изменения активного центра карбоксипептидазы А
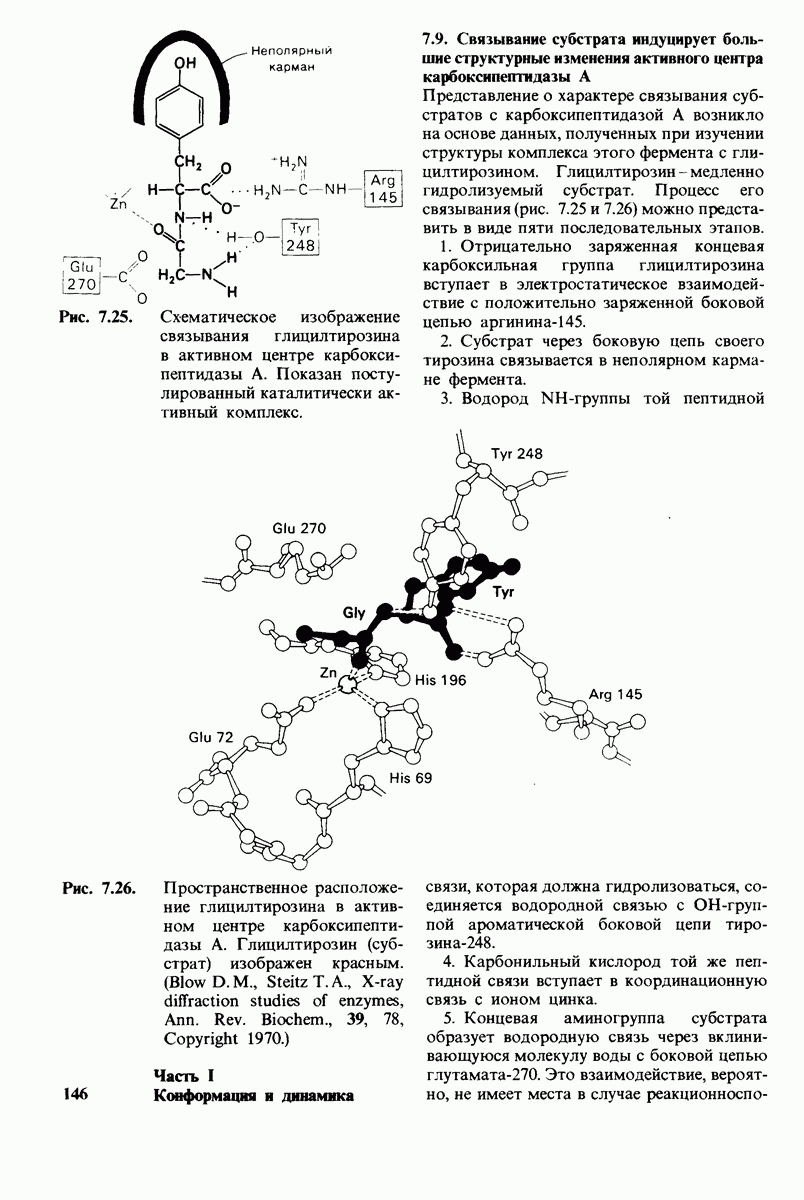
Представление о характере связывания субстратов с карбоксипептидазой А возникло на основе данных, полученных при изучении структуры комплекса этого фермента с глицилтирозином. Глицилтирозин - медленно гидролизуемый субстрат. Процесс его связывания (рис. 7.25 и 7.26) можно представить в виде пяти последовательных этапов.
1. Отрицательно заряженная концевая карбоксильная группа глицилтирозина вступает в электростатическое взаимодействие с положительно заряженной боковой цепью аргинина-145.
2. Субстрат через боковую цепь своего тирозина связывается в неполярном кармане фермента.
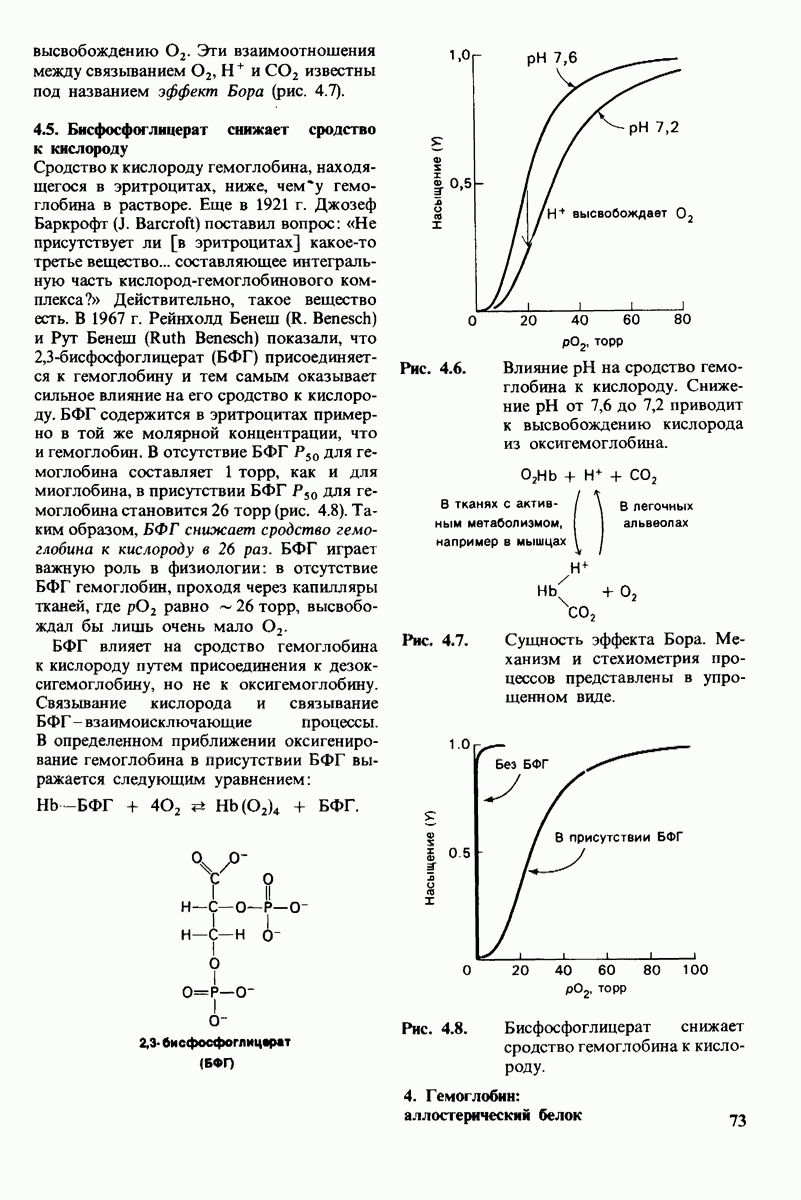
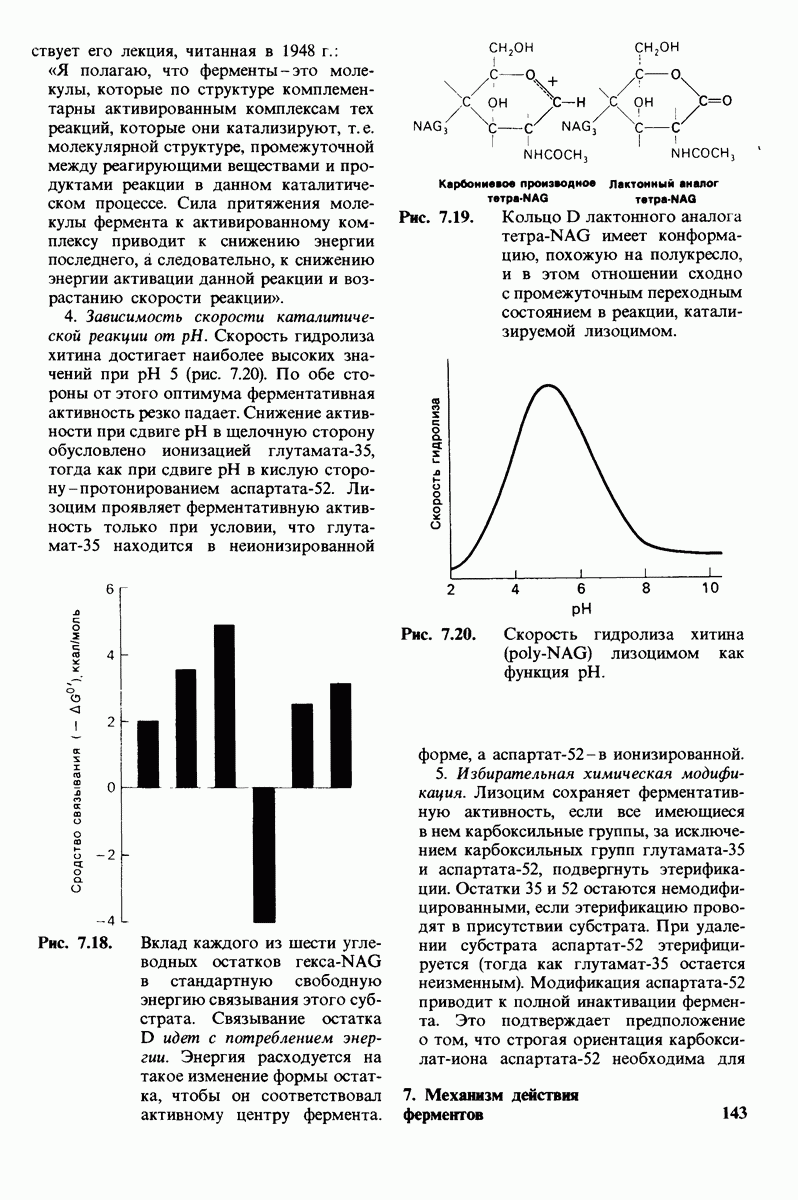
3. Водород NH-группы той пептидной связи, которая должна гидролизоваться, соединяется водородной связью с ОН-группой ароматической боковой цепи тирозина-248.
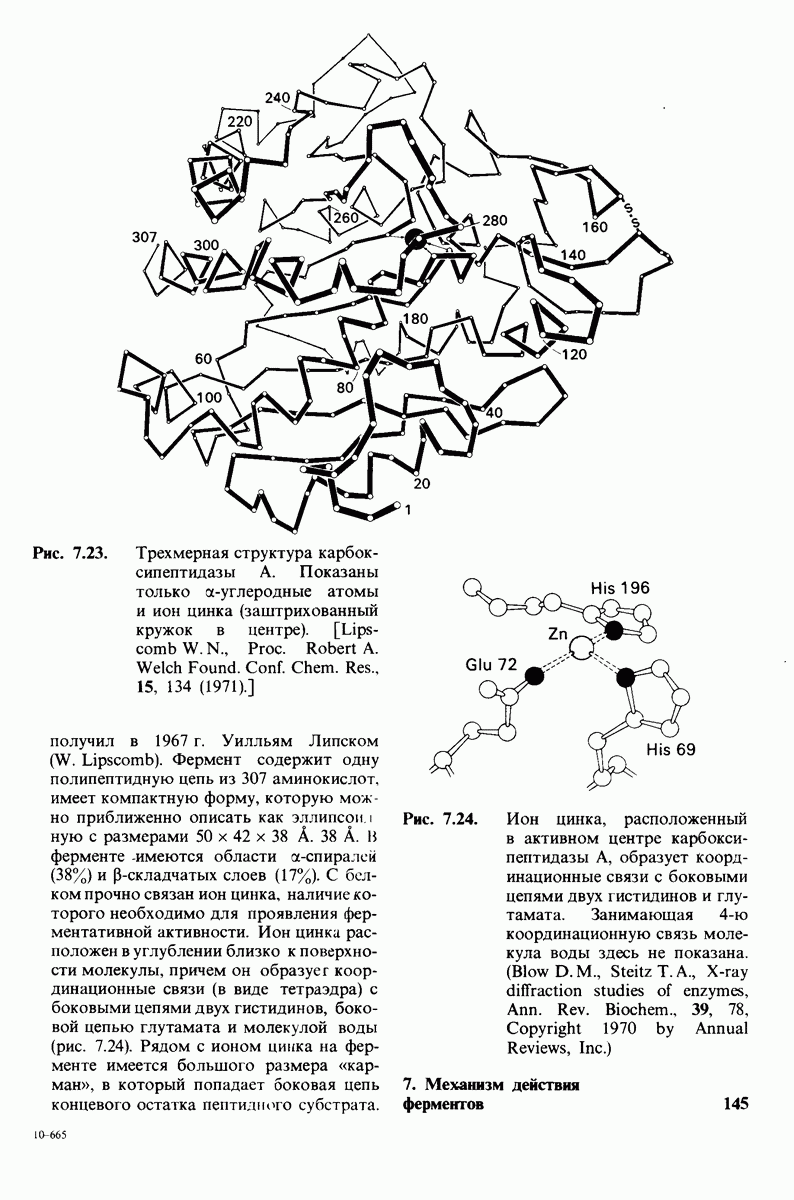
4. Карбонильный кислород той же пептидной связи вступает в координационную связь с ионом цинка.
5. Концевая аминогруппа субстрата образует водородную связь через вклинивающуюся молекулу воды с боковой цепью глутамата-270. Это взаимодействие, вероятно, не имеет места в случае

(кликните для просмотра скана)

Рис. 7.28.
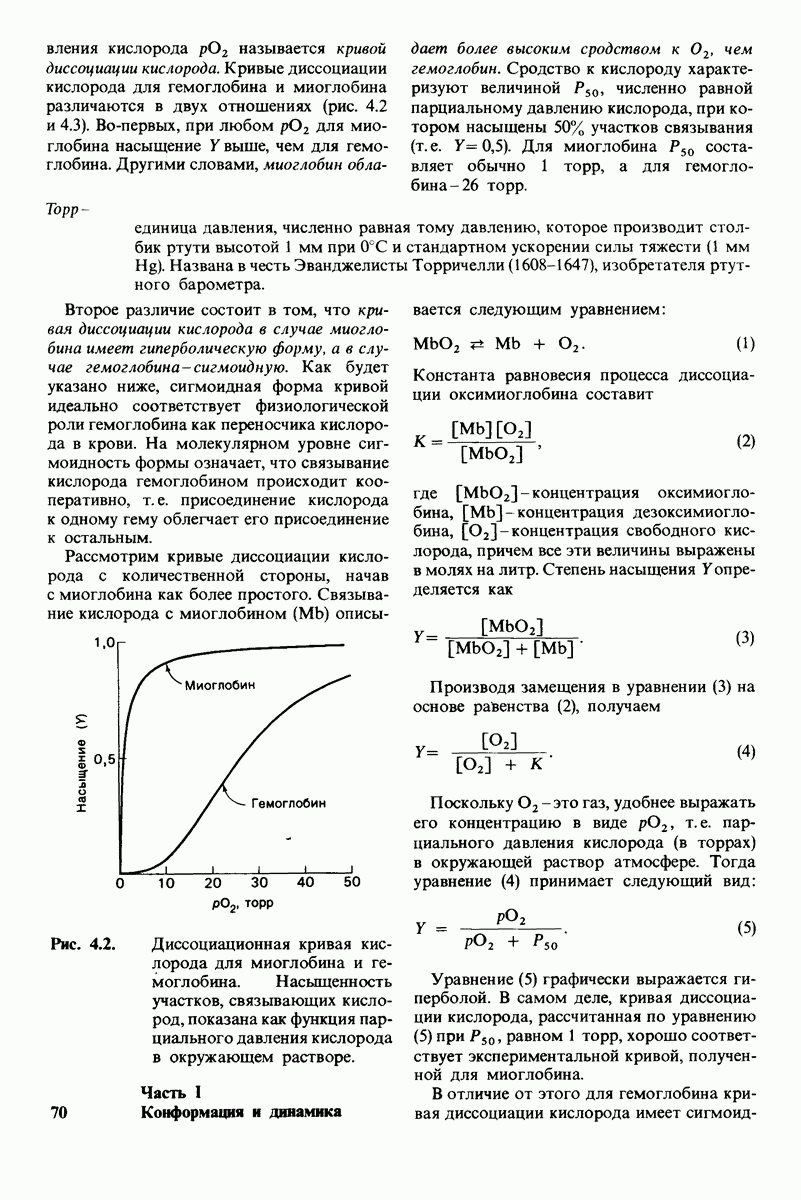
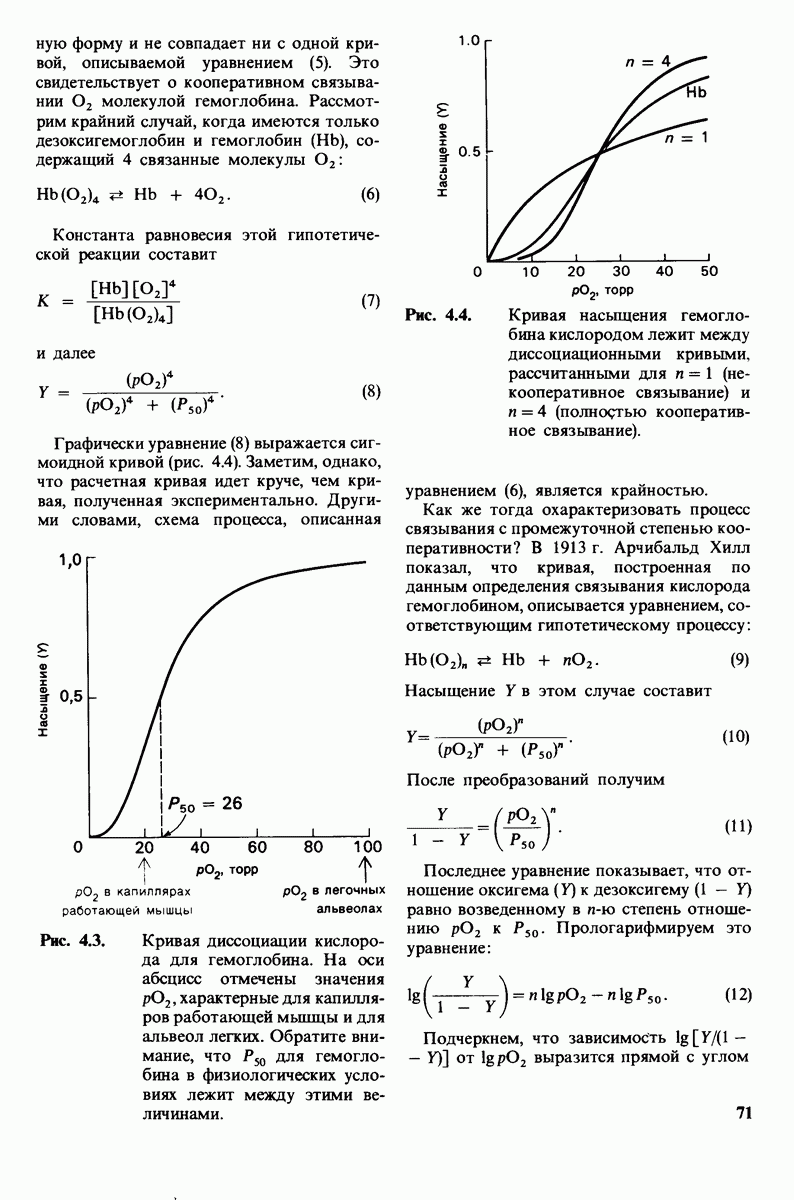
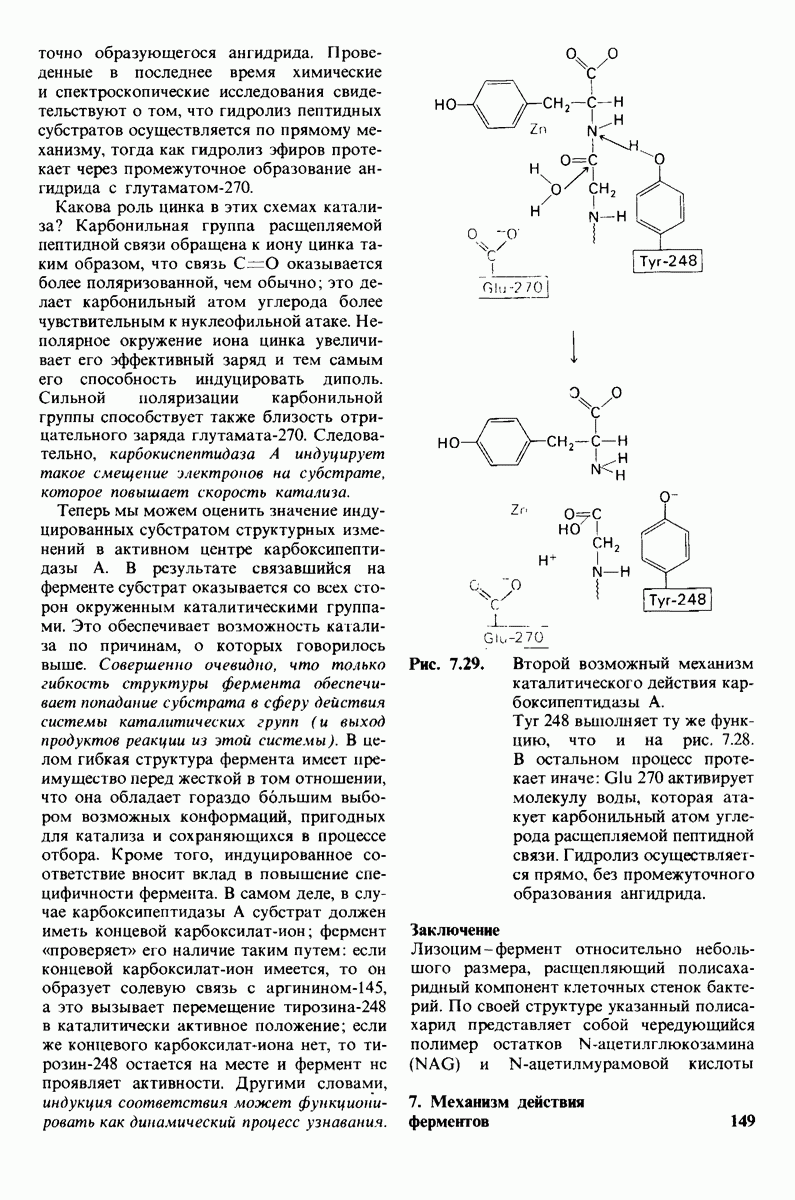
Постулированный механизм каталитического действия кар-боксипептидазы  непосредственно атакует карбонильный углеродный атом ги-дролизуемой пептидной связи, а
непосредственно атакует карбонильный углеродный атом ги-дролизуемой пептидной связи, а  отдает протон на NH-группу этого пептида. Образующийся в результате ангидрид далее подвергается гидролизу.
отдает протон на NH-группу этого пептида. Образующийся в результате ангидрид далее подвергается гидролизу.
реакционноспособного  -комплекса, и именно оно, возможно, является причиной крайне медленного гидролиза глицилтирозина.
-комплекса, и именно оно, возможно, является причиной крайне медленного гидролиза глицилтирозина.
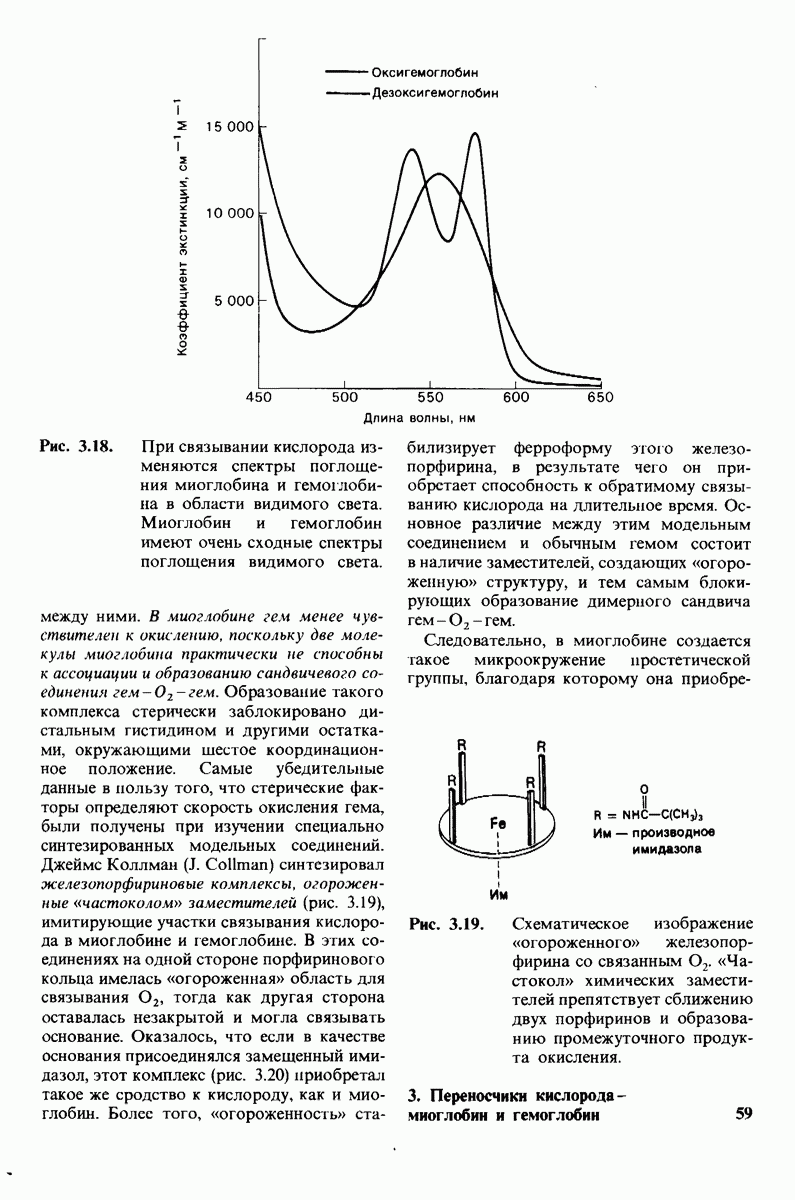
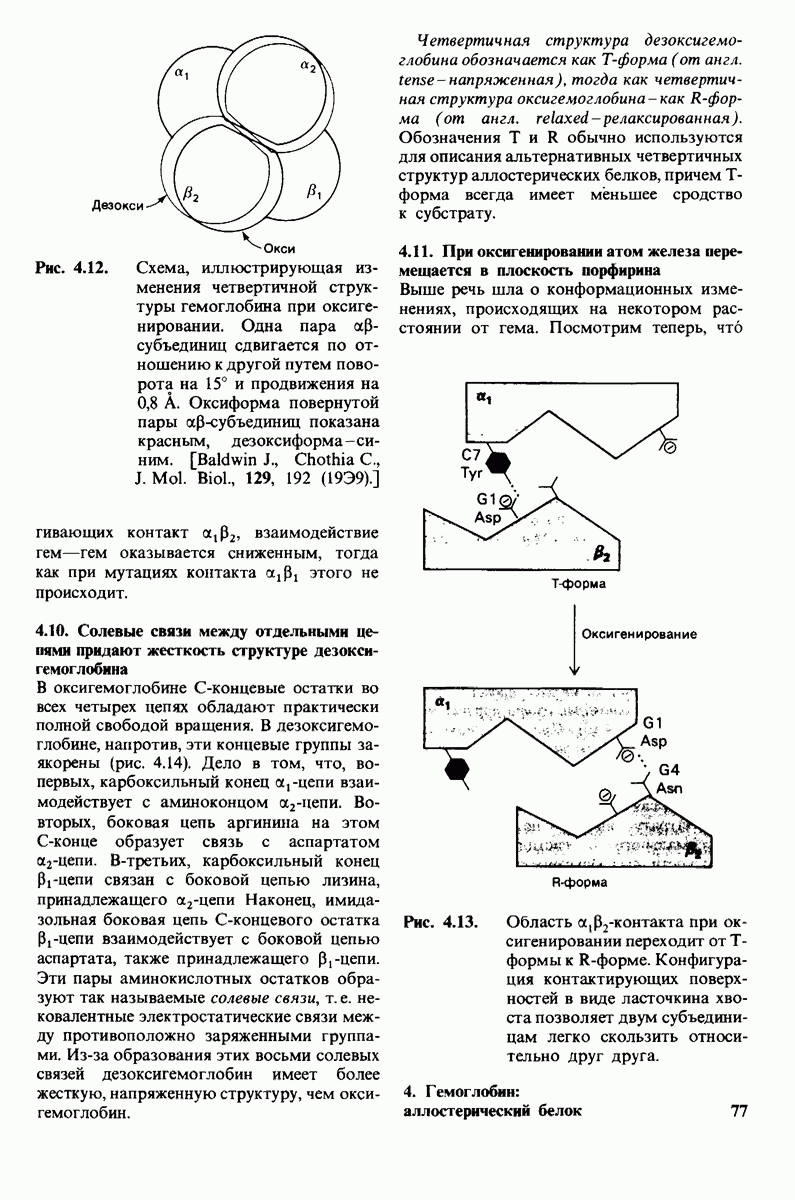
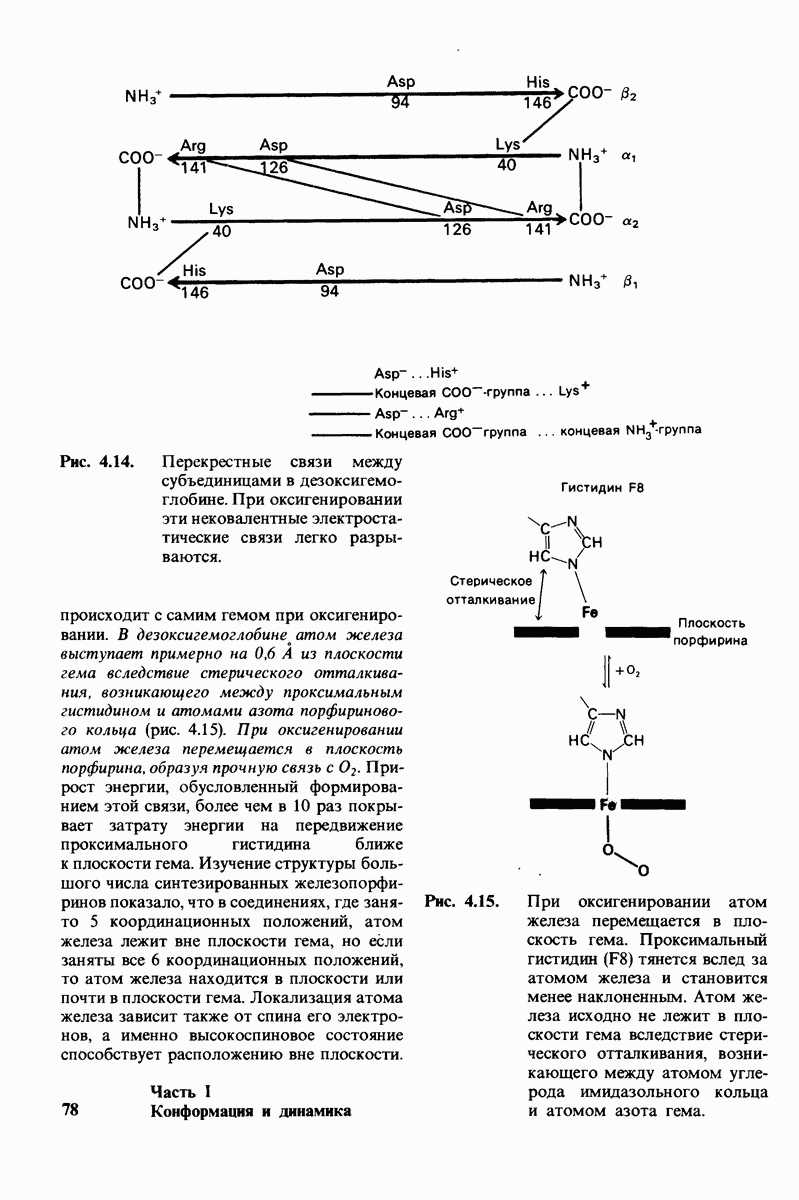
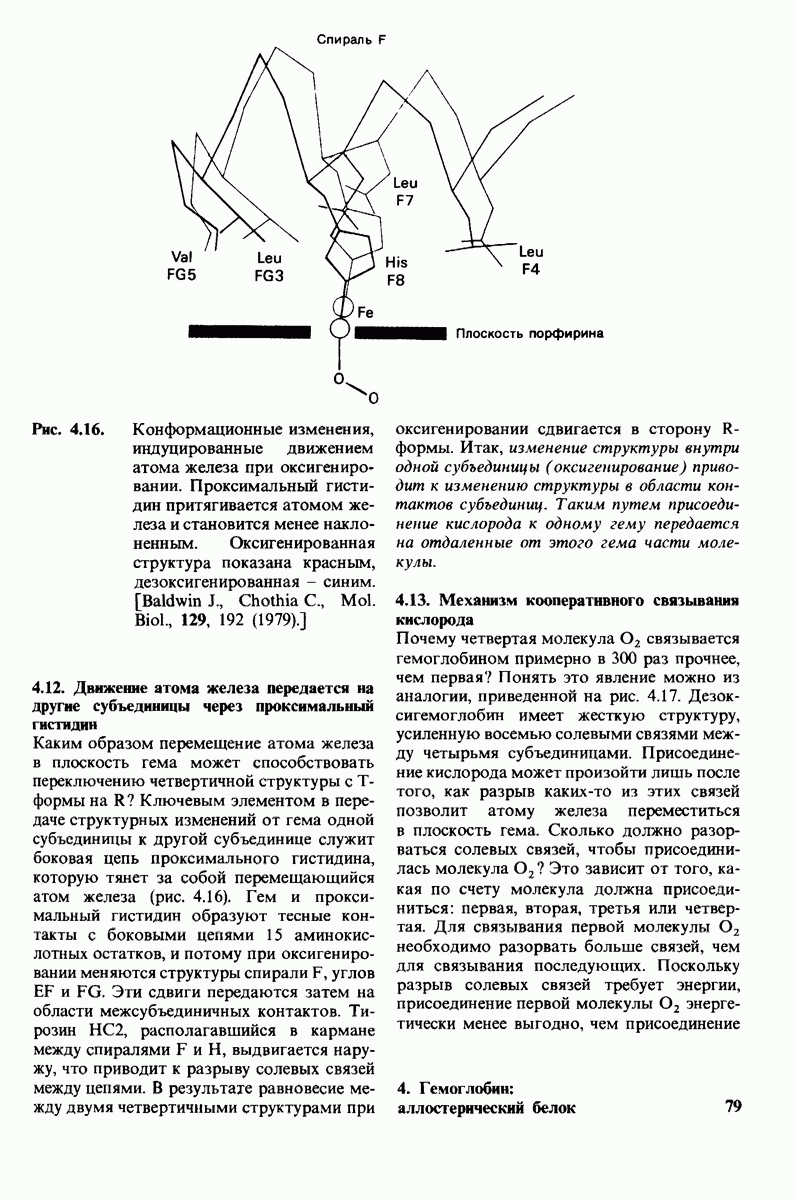
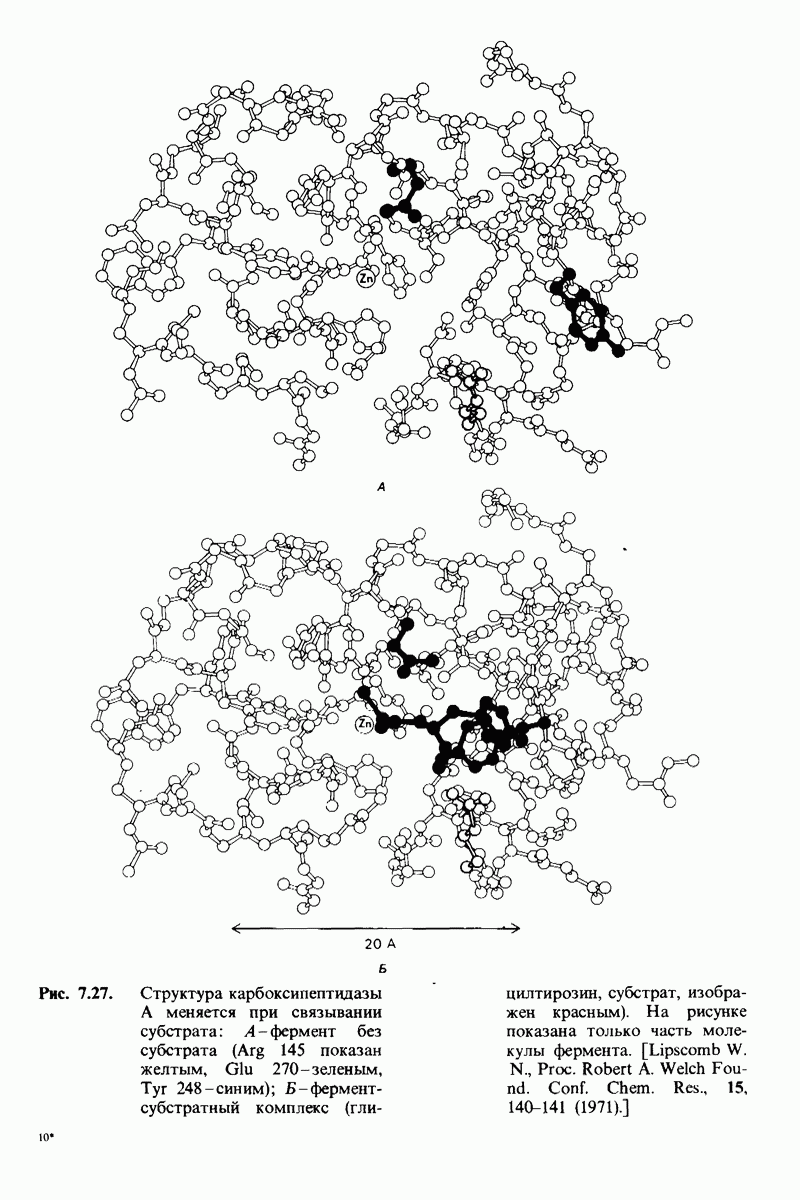
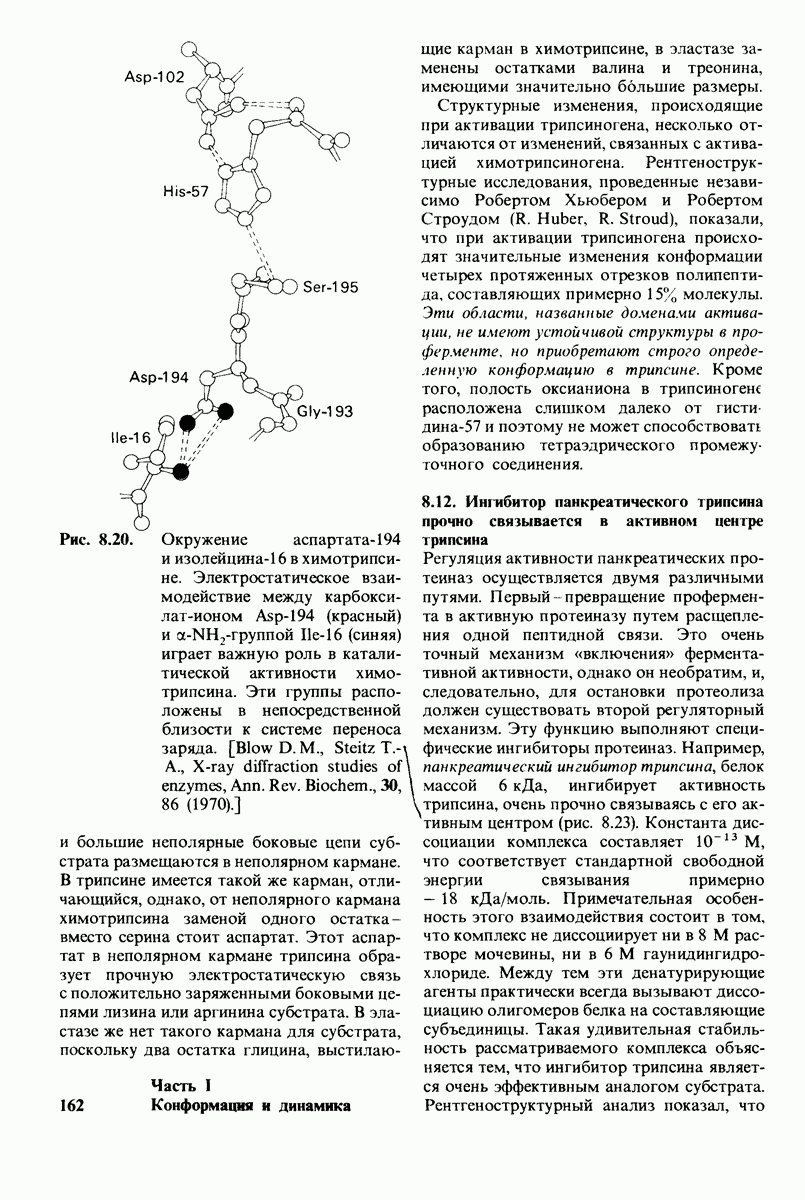
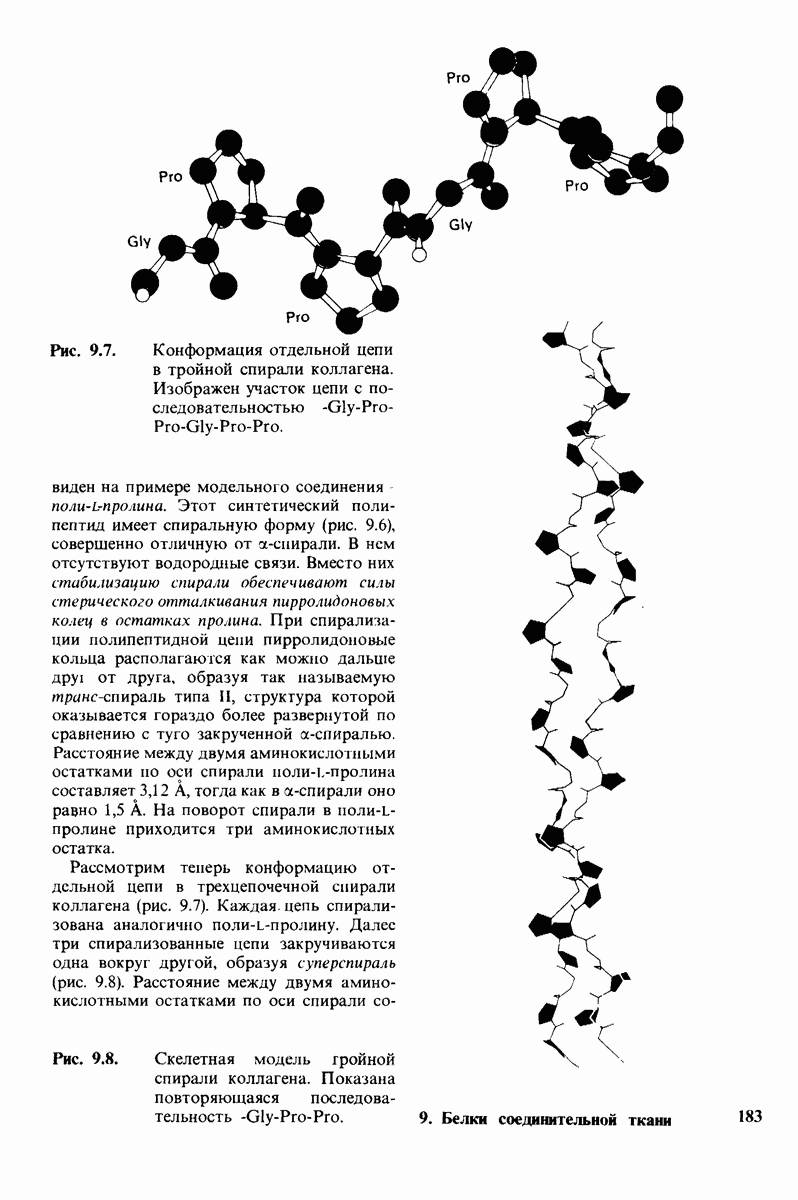
Связывание глицилтирозина сопровождается структурной перестройкой активного центра (рис. 7.27). В сущности, только присоединив субстрат, каталитические группы фермента принимают правильную ориентацию - положение, впервые постулированное Кошландом (Koshland) в его модели индуцированного соответствия. Гуанидиниевая группа аргинина-145, а также карбоксильная группа глутамата-270 перемещаются на 2 А. Связывание карбонильной группы субстрата с ионом цинка вытесняет из связи с цинком молекулу воды. По крайней мере еще четыре молекулы воды вытесняются из неполярного кармана на ферменте при связывании с ними тирозиновой боковой цепи в молекуле субстрата. Самое большое изменение конформации - это перемещение фенольного гидроксила тирозина-248 на 12 А, т. е. на расстояние, составляющее около  диаметра молекулы. Такое перемещение осуществляется в первую очередь, путем свободного вращения относительно одинарной связи
диаметра молекулы. Такое перемещение осуществляется в первую очередь, путем свободного вращения относительно одинарной связи  и состоит в том, что гидроксильная группа тирозина-248, бывшая на поверхности молекулы, перемещается, оказываясь вблизи пептидной связи субстрата. В результате закрывается полость активного центра и тем самым завершается превращение ее из области, заполненной водой, в гидрофобную. Все эти структурные изменения инициируются, по-видимому, связыванием аргинина-145 с концевой карбоксильной группой субстрата.
и состоит в том, что гидроксильная группа тирозина-248, бывшая на поверхности молекулы, перемещается, оказываясь вблизи пептидной связи субстрата. В результате закрывается полость активного центра и тем самым завершается превращение ее из области, заполненной водой, в гидрофобную. Все эти структурные изменения инициируются, по-видимому, связыванием аргинина-145 с концевой карбоксильной группой субстрата.