Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
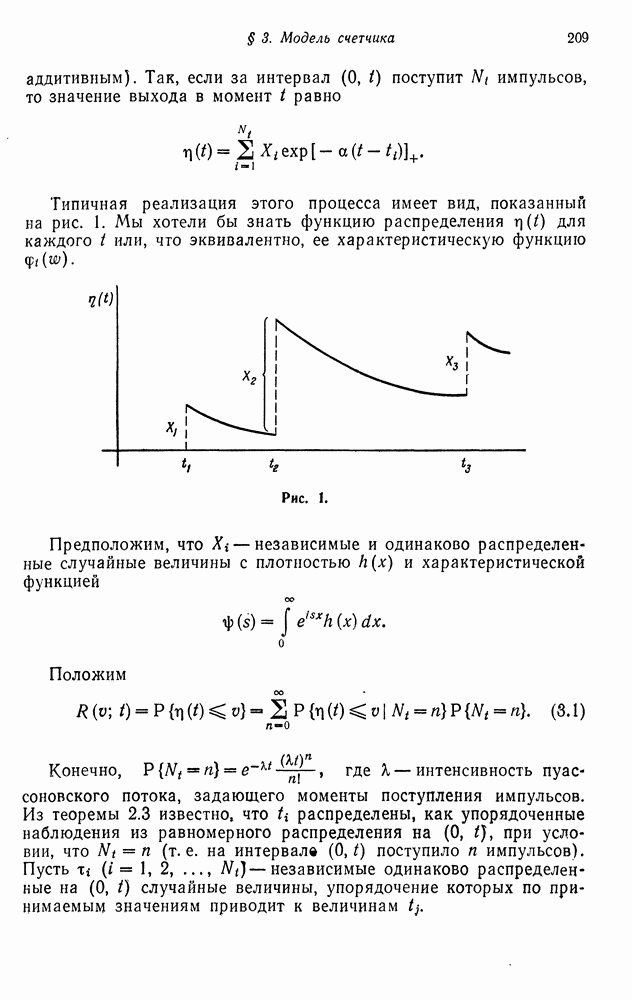
Глава 13. ДЕТЕРМИНИРОВАННЫЕ И СЛУЧАЙНЫЕ ГЕНЕТИЧЕСКИЕ И ЭКОЛОГИЧЕСКИЕ ПРОЦЕССЫОдной из главных областей применения вероятностных идей являются системы, изучаемые в генетике. В § 1 мы приведем качественное описание основных понятий, лежащих в основе генетического механизма, а в следующих двух параграфах сконцентрируем наше внимание на некоторых классических детерминированных моделях. В § 4 будет рассмотрено несколько вероятностных моделей генетических систем, а также будет обсуждена их взаимосвязь с ветвящимися процессами. Представленный ниже материал следует расценивать как введение в богатый и чрезвычайно интересный раздел генетики популяции, связанный с ее внутренней вероятностной структурой. § 1. ГЕНЕТИЧЕСКИЕ МОДЕЛИ. ОПИСАНИЕ ГЕНЕТИЧЕСКОГО МЕХАНИЗМАВ ядре клетки (рис. 1) находятся хромосомы, которые «ответственны» за наследственные характеристики организма. Их число различно для различных организмов. Единица, контролирующая полностью или частично наследование некоторого признака, называется геном. Место, которое ген занимает в хромосоме, называется локусом. Набор хромосом может содержать по одной хромосоме каждого типа (гаплоиды), как у некоторых грибов, по паре хромосом (ди-плоиды), как у млекопитающих, или группы большего размера (триплоиды, тетраплоиды и в общем случае полиплоиды), как у многих растений. В каждом локусе гены могут принадлежать к различным контрастирующим типам, которые называются аллелями. Различные возможные комбинации аллелей в соответствующих локусах разных хромосом называются генотипами. При диплоидии, если имеются аллели А и а, возможны следующие генотипы:
Фенотипами называются различные внешние проявления аллельных комбинаций.
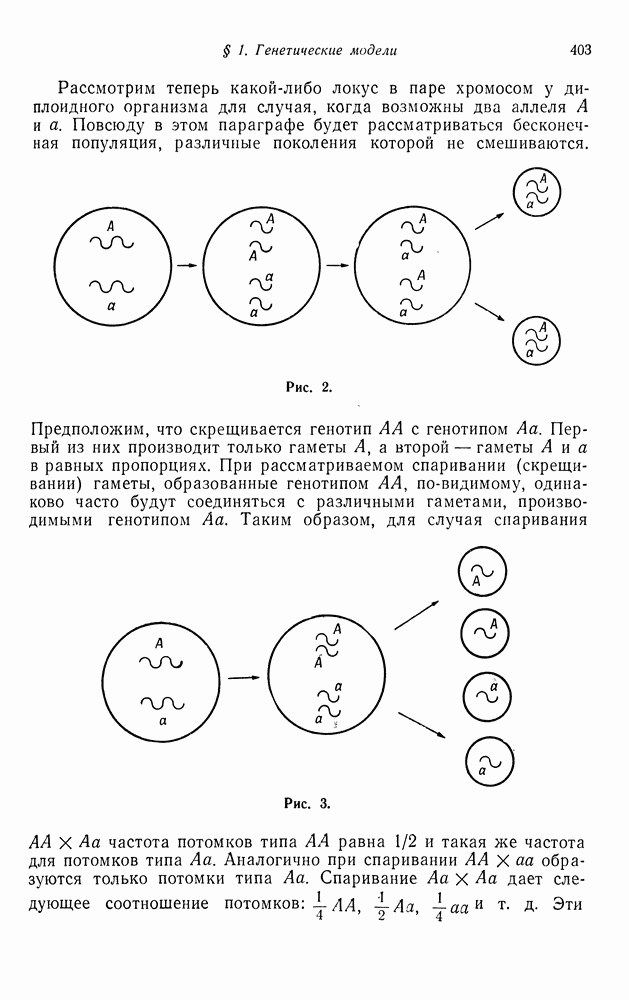
Рис. 1. Например, если А доминирует над а, то разные генотипы А. Размножение и закон Харди — ВейнбергаСуществует два способа клеточного деления. Более распространенным из них является митоз (рис. 2). При этом способе каждая хромосома делится на две и, когда клетка расщепляется на две дочерние клетки, каждая из вновь образованных клеток получает по одной хромосоме из каждой пары. Таким образом, каждая новая клетка имеет полный набор хромосом. Процесс второго типа — мейоз (рис. 3) — происходит при половом размножении диплоидных клеток. Каждая хромосома при этом удваивается. Затем образуются четыре новые клетки, каждая из которых содержит одну хромосому. Таким образом, каждая из четырех новых клеток содержит половину нормального набора хромосом. Такая клетка называется гаметой. Когда две гаметы соединяются, образуется оплодотворенная клетка — зигота, которая имеет полный набор хромосом. Рассмотрим теперь какой-либо локус в паре хромосом у диплоидного организма для случая, когда возможны два аллеля А и а. Повсюду в этом параграфе будет рассматриваться бесконечная популяция, различные поколения которой не смешиваются.
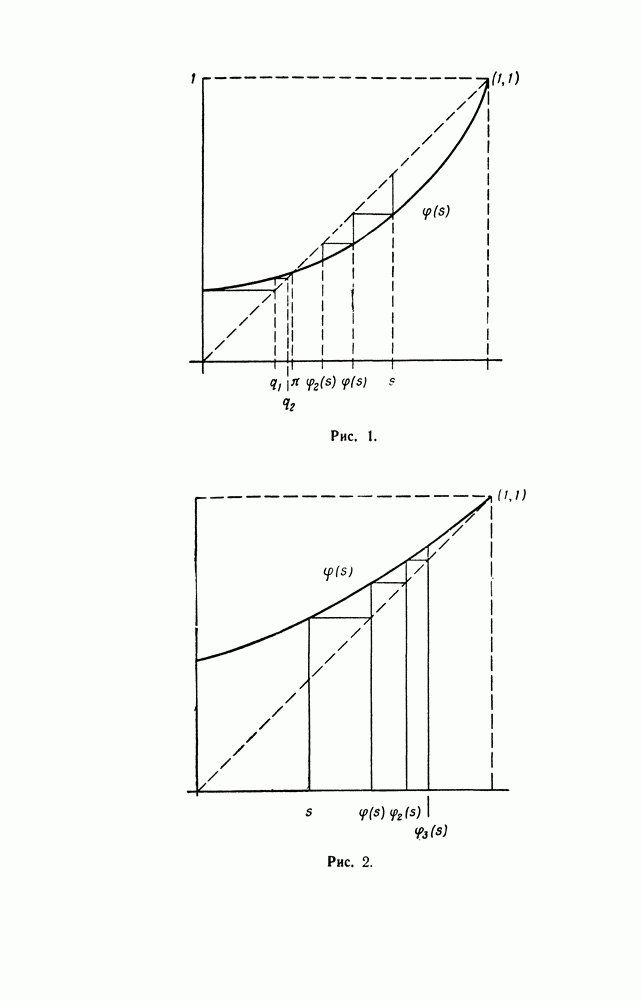
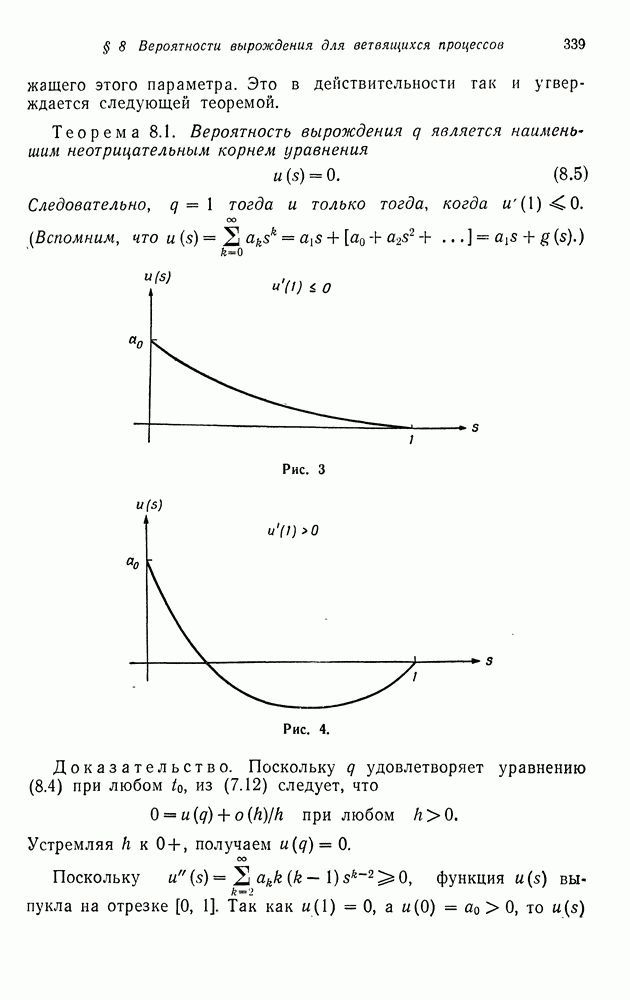
Рис. 2. Предположим, что скрещивается генотип
Рис. 3. Таким образом, для случая спаривания пропорции исходов различных возможных спариваний известны под названием отношения при расщеплении. Рассмотрим некоторую раздельнополую популяцию с тремя генотипами Таблица I
Следовательно, в следующем поколении эти три генотипа будут представлены в отношении
Положим
и р + q = 1. Точно таким же образом мы найдем, что после следующего спаривания генотипы распределяются в отношении
Таким образом, начиная со второго поколения, соотношение между генотипами не меняется, т. е. устойчивое состояние достигается за одно поколение. Этот результат известен, как закон Харди — Вейнберга. Можно получить этот же результат следующим образом. Рассмотрим популяцию гамет: если начальное соотношение между генотипами то же, что и выше, то соответствующая доля гамет А в популяции равна
Рассматривая совокупность гамет в новой популяции, найдем:
что указывает на достигнутую устойчивость. Факт совпадения результатов, полученных обоими методами, известен в виде следующего предложения: «случайное слияние гамет» эквивалентно случайному спариванию. (Вероятностная интерпретация этой теоремы содержится в задаче 17.) Теперь мы рассмотрим, как три фактора — мутация, миграция и отбор — влияют на популяцию генотипов в предположении о случайном спаривании. Б. Мутация1. Рассмотрим гаплоидную популяцию генов А и а. Предположим, что в промежутке между любыми двумя последующими поколениями доля 2. Пусть теперь между двумя последующими поколениями наряду с рассмотренной мутацией доля
что эквивалентно соотношению
По индукции получаем
Снова сходимость имеет геометрический характер. Заметим, что если В. МиграцияПредположим, что мы имеем 2 популяции, составленные из генов Тогда Г. ОтборСуществует 2 вида отбора — гаметический, который отражает относительные преимущества некоторых гамет над другими, и зиготный, который отражает преимущества некоторых генотипов над другими. Такие преимущества могут соответствовать различным смертностям, относительной плодовитости и т. д. (а) Гаметический отбор Пусть Таблица II
Сформируем следующее поколение гамет
Сомножитель
Отсюда
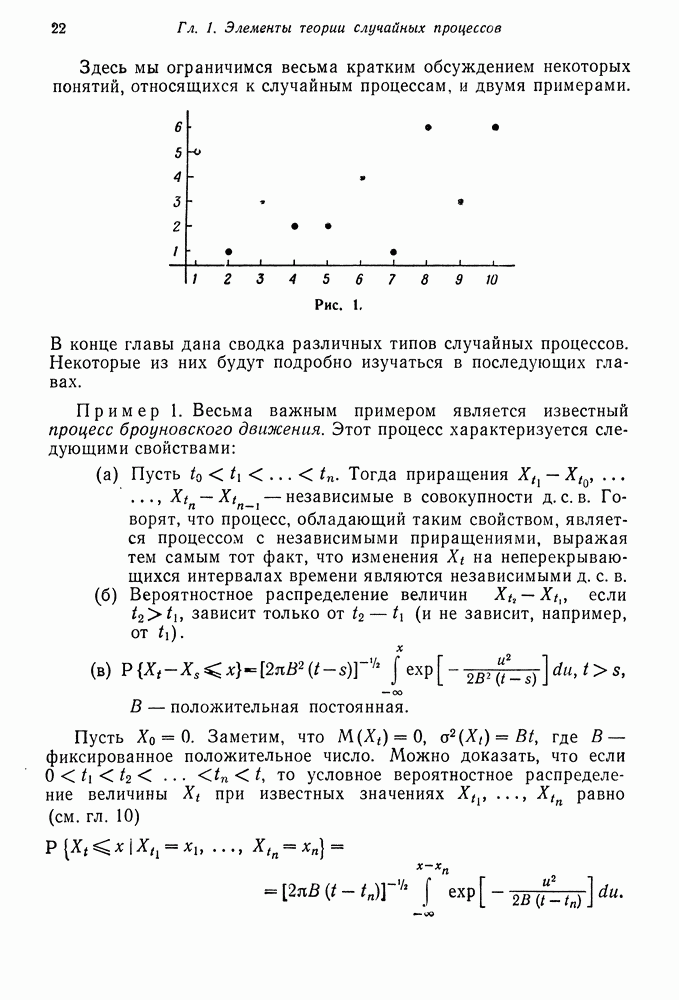
Следовательно, (б) Зиготный отбор. Общий случай Рассмотрим диплоидную популяцию, в которой каждый генотип характеризуется индексом Таблица III
Пусть Зиготные частоты
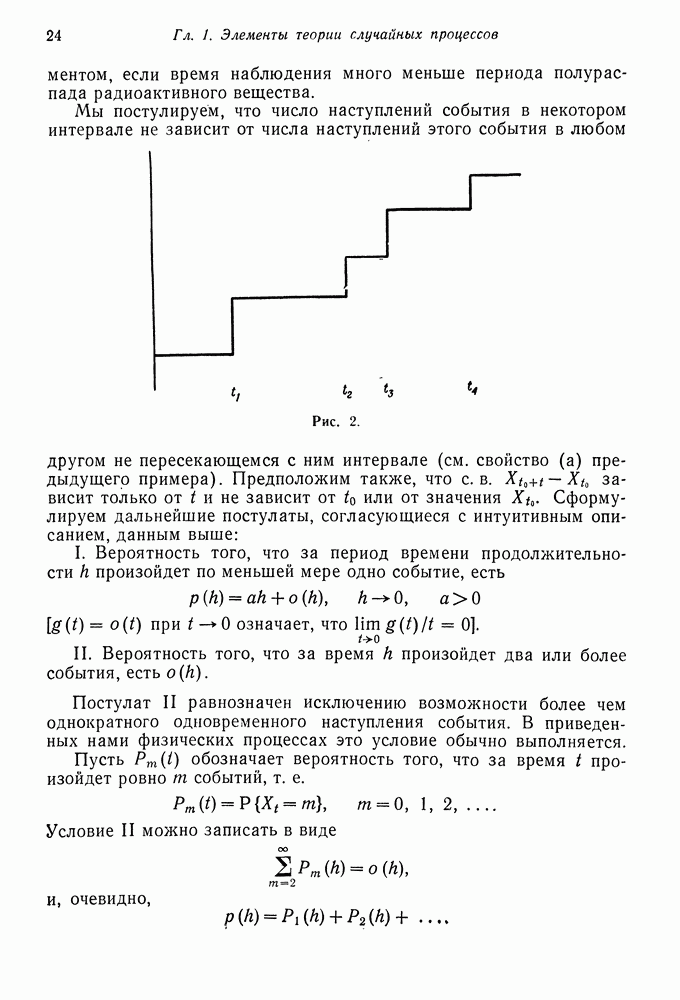
где величина
Таким образом,
Равновесной частотой является любая величина каждый генотип. Такая ситуация имеет место, когда гетерозиготный индивидуум имеет селективное преимущество перед каждым гомозиготным индивидуумом, как, например, при наследовании признака серповидноклеточной анемии. Пример полиморфизма: (а) (б)
Пусть Д. Модели двуполых популяцийРассмотрим теперь популяцию, состоящую из мужских и женских особей, спаривание при этом происходит только между представителями различных полов. Если ограничиться рассмотрением лишь генов, локализованных на аутосомных (неполовых) хромосомах (как это было в предыдущем случае), то процесс случайного спаривания можно описать точно таким же образом, как и выше. Предположим, что частоты генотипов внутри мужской и женской популяций равны:
Гаметы имеют следующие частоты:
Для случайного спаривания, которое эквивалентно случайному выбору из мужских и женских совокупностей гамет, мы найдем, что частоты генотипов в обеих мужской и женской популяциях во втором поколении равны Однако, если рассмотреть половые хромосомы, ситуация резко изменится. Причина этого состоит в том, что в некоторых диплоидных клетках могут находиться два различных типа хромосом Рассмотрим распределение сцепленного с полом гена в последовательных поколениях. Предположим, что начальные частоты генотипов у мужских и женских особей равны
Заметим, что генотип у мужской особи определяется только одним символом гена, поскольку Y-хромосома не содержит ни одного гена, содержащегося в Х-хромосоме. Частоты гамет у женских особей равны
В предположении о случайном спаривании, или, что то же, о случайном выборе из совокупности гамет, частоты генотипов в следующем поколении равны у женских особей:
и у мужских особей
Каждая вновь образованная мужская зигота получает Y-xpoмосому из мужской популяции и Х-хромосому из женской. Следовательно, доля генов А в мужской популяции в точности равна доле генов А в предыдущем поколении женской популяции. В общем случае если
Отсюда можно вывести рекуррентное соотношение
которое сводится к следующему:
или
или
Отсюда
В пределе
получили выражение для предельной частоты генов А в мужской популяции. В женской популяции предельные частоты генотипов имеют вид
Это, конечно, в точности соответствует частотам Харди-Вейнберга.
|
 |
Оглавление
|





















































































































































































































































































































































































































































































































 Генотипы
Генотипы  и
и  называются гомозиготными, а генотип
называются гомозиготными, а генотип  гетерозиготным.
гетерозиготным.

 и
и  будут определять один и тот же фенотип, а комбинация
будут определять один и тот же фенотип, а комбинация  определяет другой фенотип.
определяет другой фенотип.

 с генотипом
с генотипом  Первый из них производит только гаметы
Первый из них производит только гаметы  а второй — гаметы А и а в равных пропорциях. При рассматриваемом спаривании (скрещивании) гаметы, образованные генотипом
а второй — гаметы А и а в равных пропорциях. При рассматриваемом спаривании (скрещивании) гаметы, образованные генотипом  по-видимому, одинаково часто будут соединяться с различными гаметами, производимыми генотипом
по-видимому, одинаково часто будут соединяться с различными гаметами, производимыми генотипом 

 частота потомков типа
частота потомков типа  равна 1/2 и такая же частота для потомков типа
равна 1/2 и такая же частота для потомков типа  Аналогично при спаривании
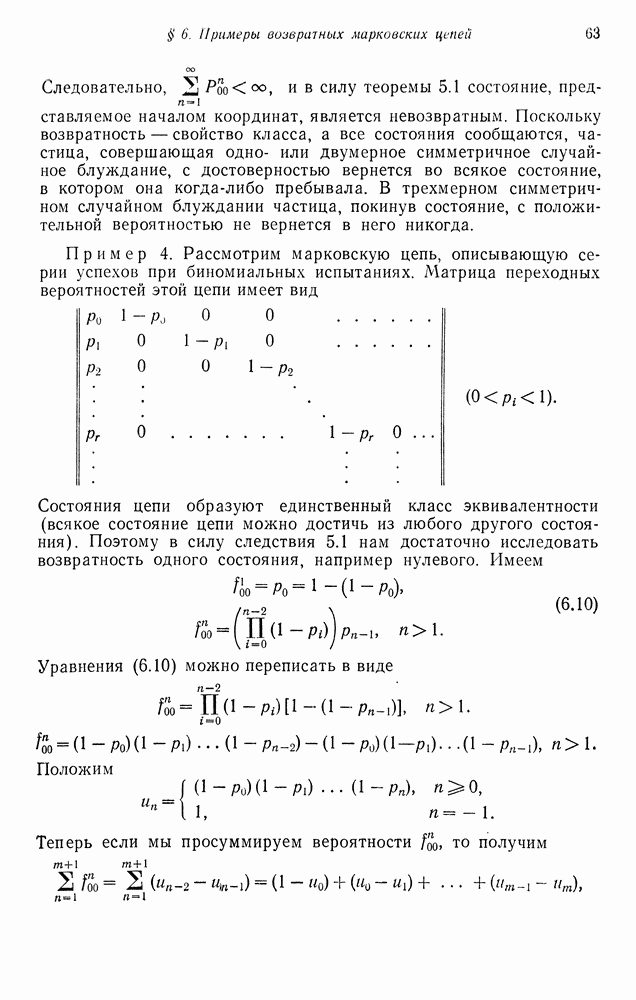
Аналогично при спаривании  образуются только потомки типа
образуются только потомки типа  Спаривание
Спаривание  дает следующее соотношение потомков:
дает следующее соотношение потомков:  Эти
Эти
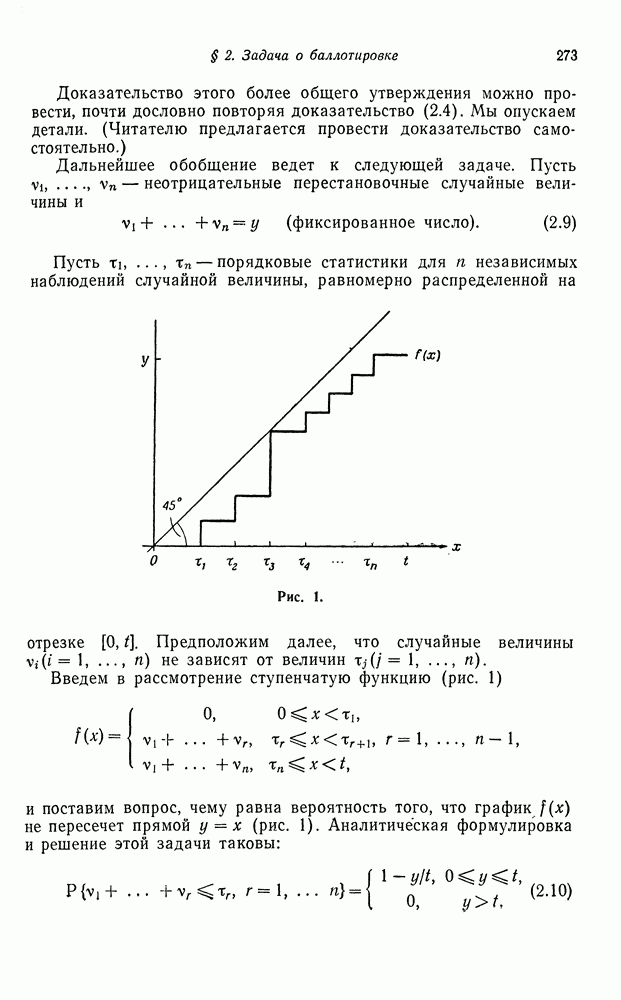
 в которой осуществляется случайное спаривание. Пусть исходное соотношение генотипов следующее: доля
в которой осуществляется случайное спаривание. Пусть исходное соотношение генотипов следующее: доля  равна
равна  доля
доля  равна
равна  и доля
и доля  равна
равна  . В табл. I показаны результаты случайного спаривания.
. В табл. I показаны результаты случайного спаривания.


 тогда эти пропорции примут вид
тогда эти пропорции примут вид


 а гамет а равна до
а гамет а равна до  так что в следующем поколении генотипы представлены в отношении
так что в следующем поколении генотипы представлены в отношении


 генов а мутирует (изменяется) в форму А. Если
генов а мутирует (изменяется) в форму А. Если  доля генов а в
доля генов а в  поколении, то
поколении, то  доля генов А в
доля генов А в  поколении. Тогда
поколении. Тогда  и окончательно
и окончательно  т. е. если
т. е. если  то 0 по геометрической прогрессии со знаменателем
то 0 по геометрической прогрессии со знаменателем 
 генов А мутирует в форму а. Тогда
генов А мутирует в форму а. Тогда



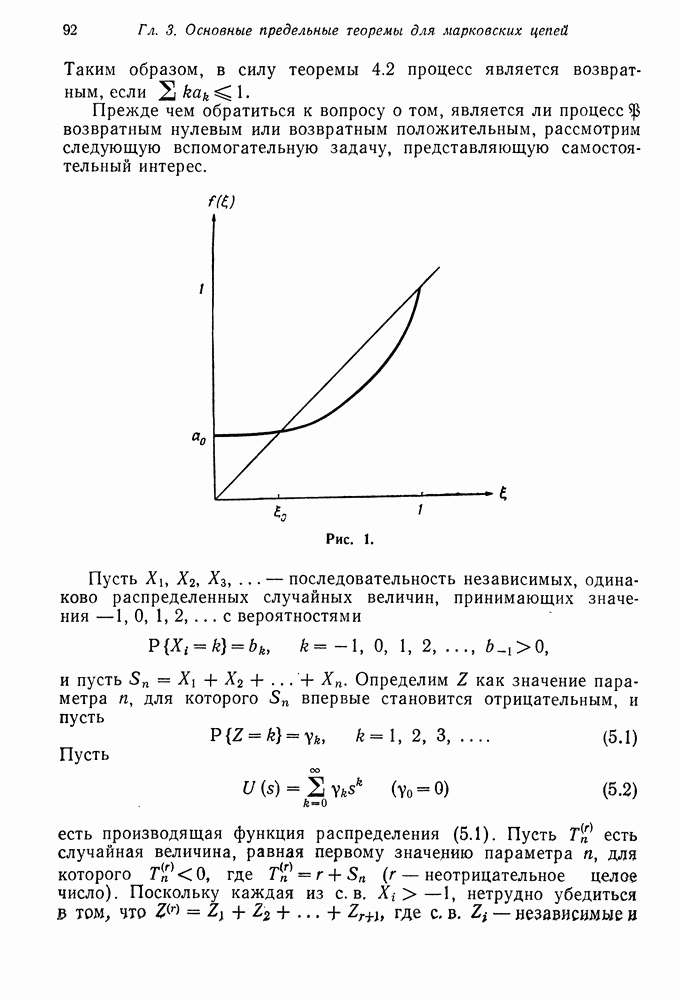
 то
то  будет сходиться к пределу немонотонно, если же
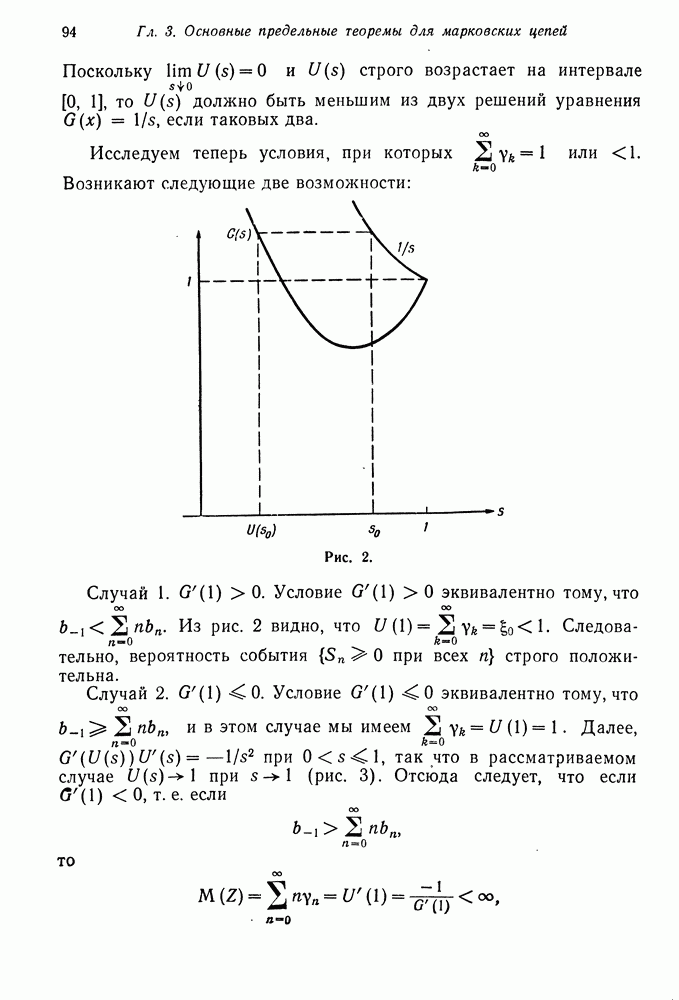
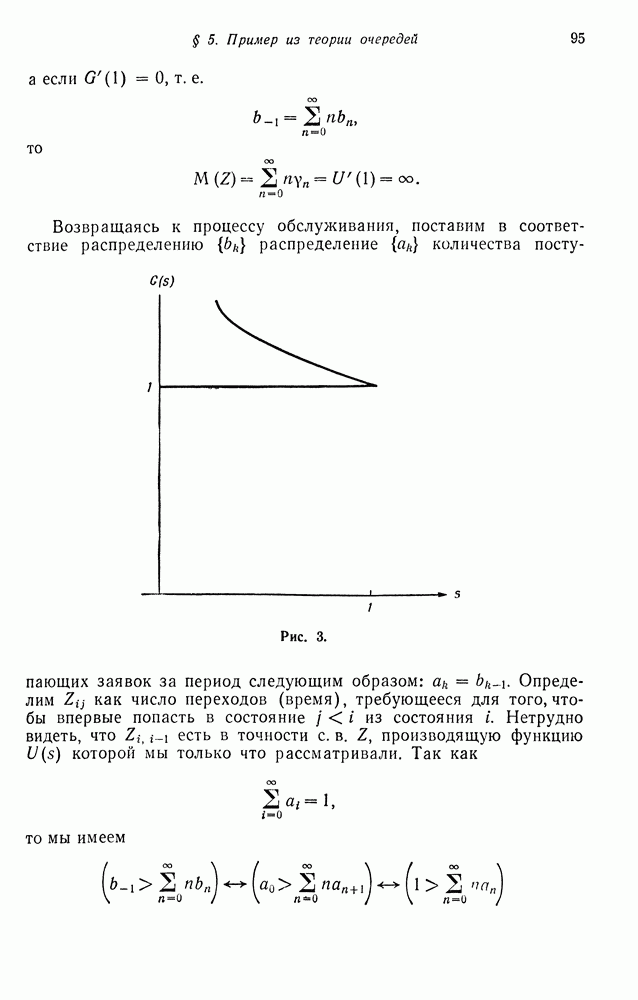
будет сходиться к пределу немонотонно, если же  то сходимость монотонная
то сходимость монотонная
 внутреннюю популяцию, в которой частоты этих генов равны
внутреннюю популяцию, в которой частоты этих генов равны  соответственно (и их мы хотим найти), и внешнюю «большую» популяцию, в которой гены А и а имеют постоянные частоты
соответственно (и их мы хотим найти), и внешнюю «большую» популяцию, в которой гены А и а имеют постоянные частоты  «Миграция» означает, что доля
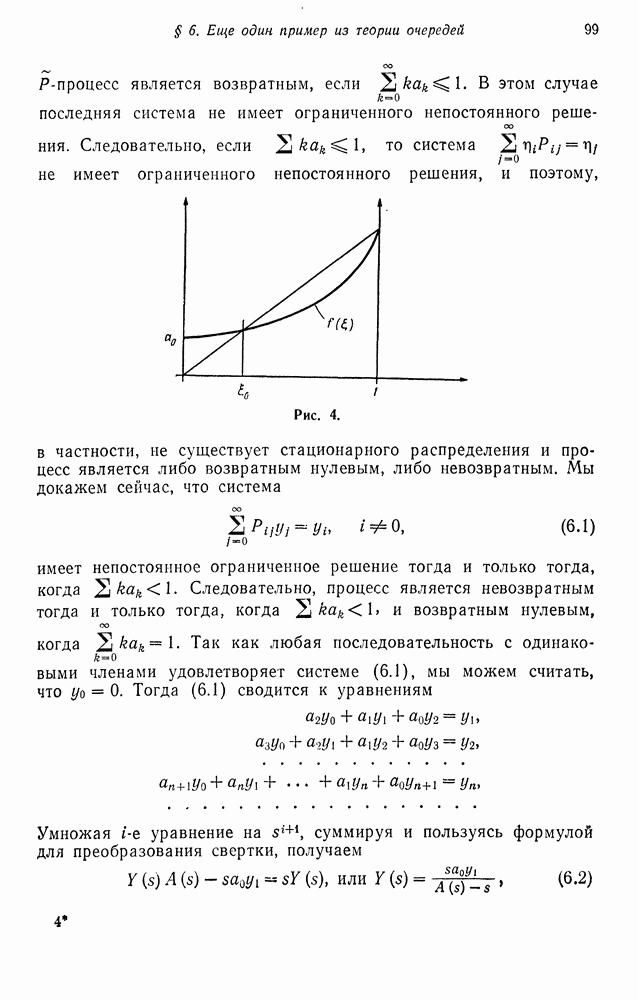
«Миграция» означает, что доля  всей внутренней популяции за время одного поколения замещается таким же количеством индивидуумов, взятых из внешней популяции.
всей внутренней популяции за время одного поколения замещается таким же количеством индивидуумов, взятых из внешней популяции.
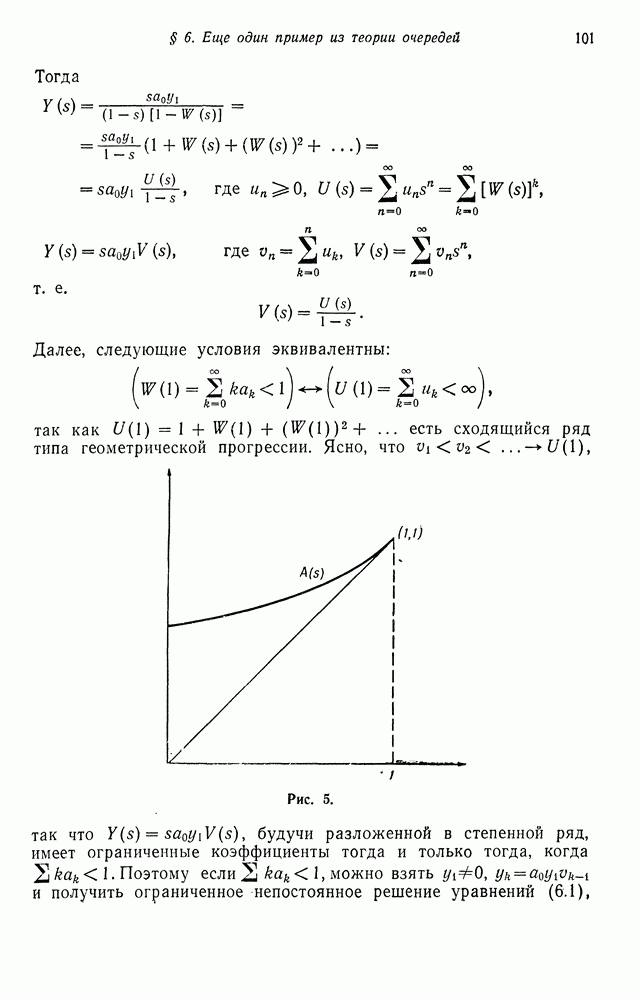
 Поскольку величина
Поскольку величина  постоянна, полученное рекуррентное соотношение совпадает, очевидно, с описывающим процесс мутации, если взять
постоянна, полученное рекуррентное соотношение совпадает, очевидно, с описывающим процесс мутации, если взять  Отсюда следует, что
Отсюда следует, что  сходятся к величине
сходятся к величине 
 меры относительного селективного преимущества гаметы А по сравнению с гаметой а. Пусть
меры относительного селективного преимущества гаметы А по сравнению с гаметой а. Пусть  означают их частоты в популяции в
означают их частоты в популяции в  поколении.
поколении.  данные сведены в таблицу II.
данные сведены в таблицу II.

 приписывая имеющимся частотам гамет веса
приписывая имеющимся частотам гамет веса  соответственно. Получим
соответственно. Получим

 является нормирующим. Имеем
является нормирующим. Имеем


 т. е. популяция генов а вырождается.
т. е. популяция генов а вырождается.
 отражающим его преимущество или недостаток по сравнению с другими типами. Предположим, что относительное преимущество не зависит от времени и частот гамет в
отражающим его преимущество или недостаток по сравнению с другими типами. Предположим, что относительное преимущество не зависит от времени и частот гамет в  поколении.
поколении.

 частоты
частоты  и а соответственно в
и а соответственно в  поколении (см. таблицу III).
поколении (см. таблицу III).
 поколении равны соответственно
поколении равны соответственно

 называется функцией приспособленности и
называется функцией приспособленности и  играет роль нормирующего множителя. Следовательно, частоты гамет А и а равны соответственно
играет роль нормирующего множителя. Следовательно, частоты гамет А и а равны соответственно


 , такая, что из равенства
, такая, что из равенства  следует
следует  Она называется устойчивой, если для значений
Она называется устойчивой, если для значений  близких к
близких к  переменная
переменная  при
при  стремится к
стремится к  Равновесная частота, не являющаяся устойчивой, называется неустойчивой. Определим полиморфизм как такое устойчивое равновесное состояние, в котором представлен
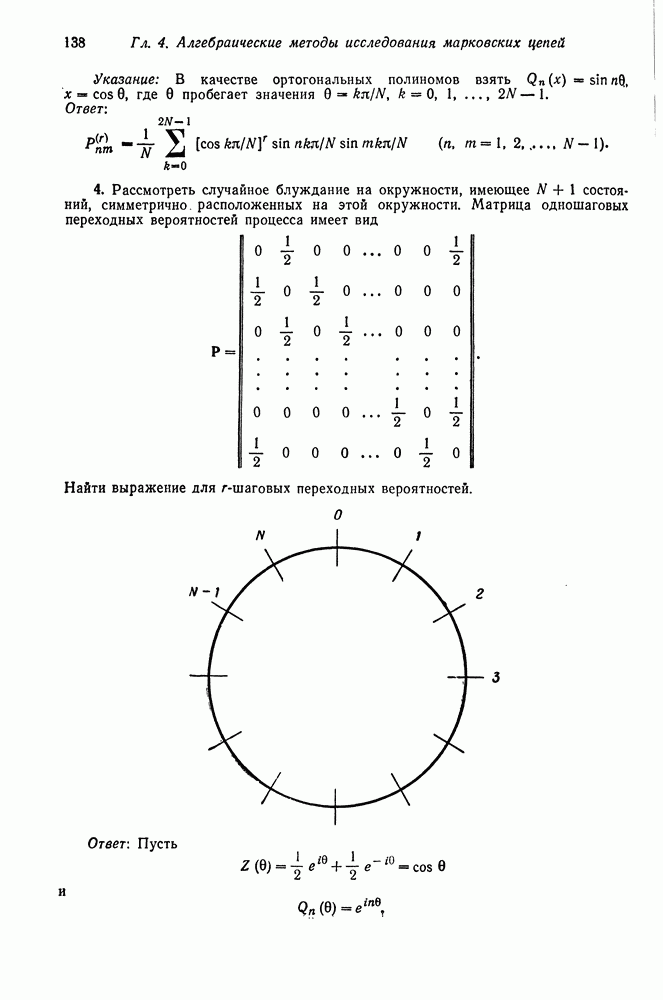
Равновесная частота, не являющаяся устойчивой, называется неустойчивой. Определим полиморфизм как такое устойчивое равновесное состояние, в котором представлен
 Такая ситуация обычно называется гетерозиготным преимуществом, или доминированием. Рассмотрим случаи, когда
Такая ситуация обычно называется гетерозиготным преимуществом, или доминированием. Рассмотрим случаи, когда  равна нулю.
равна нулю.
 . Это равновесное состояние. Однако если
. Это равновесное состояние. Однако если  близко к 1, то
близко к 1, то  и переменная
и переменная  удаляется от 1. Это неустойчивое равновесие.
удаляется от 1. Это неустойчивое равновесие.
 . Это состояние также неустойчивое, поскольку
. Это состояние также неустойчивое, поскольку  при малых
при малых 

 Тогда
Тогда  имеет знак, обратный знаку 6, так как
имеет знак, обратный знаку 6, так как  Отсюда следует, что положение равновесия устойчивое. Чуть более подробный анализ показывает, что процесс (1.1) сходится к равновесному состоянию
Отсюда следует, что положение равновесия устойчивое. Чуть более подробный анализ показывает, что процесс (1.1) сходится к равновесному состоянию  при любом начальном состоянии
при любом начальном состоянии  таком, что
таком, что 


 Для
Для  соответственно. Как и в случае самооплодотворения, устойчивое распределение достигается за одно поколение.
соответственно. Как и в случае самооплодотворения, устойчивое распределение достигается за одно поколение.
 . У млекопитающих женские особи характеризуются наличием двух гомологичных Х-хромосом, в то время как у мужских особей спарены хромосомы
. У млекопитающих женские особи характеризуются наличием двух гомологичных Х-хромосом, в то время как у мужских особей спарены хромосомы  Более того, многие, если не все, гены из Х-хромосомы не имеют комплементов на Y-хромосоме. В рассматриваемом случае генотип мужской особи полностью определяется аллелями, локализованными на Х-хромосоме. Такие гены и контролируемые ими признаки называются сцепленными с полом.
Более того, многие, если не все, гены из Х-хромосомы не имеют комплементов на Y-хромосоме. В рассматриваемом случае генотип мужской особи полностью определяется аллелями, локализованными на Х-хромосоме. Такие гены и контролируемые ими признаки называются сцепленными с полом.




 частота аллеля А в совокупности женских гамет в
частота аллеля А в совокупности женских гамет в  поколении, то
поколении, то  частота аллеля А в мужской популяции в
частота аллеля А в мужской популяции в  поколении. Следовательно,
поколении. Следовательно,  поколение женских генотипов имеет состав
поколение женских генотипов имеет состав



 где
где  . По индукции получаем
. По индукции получаем




