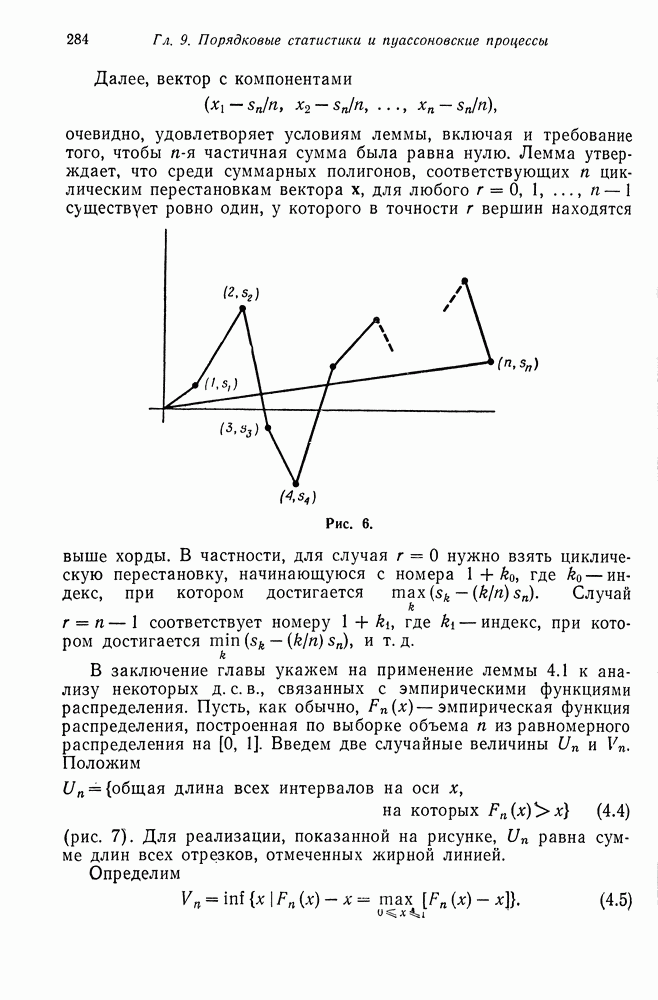
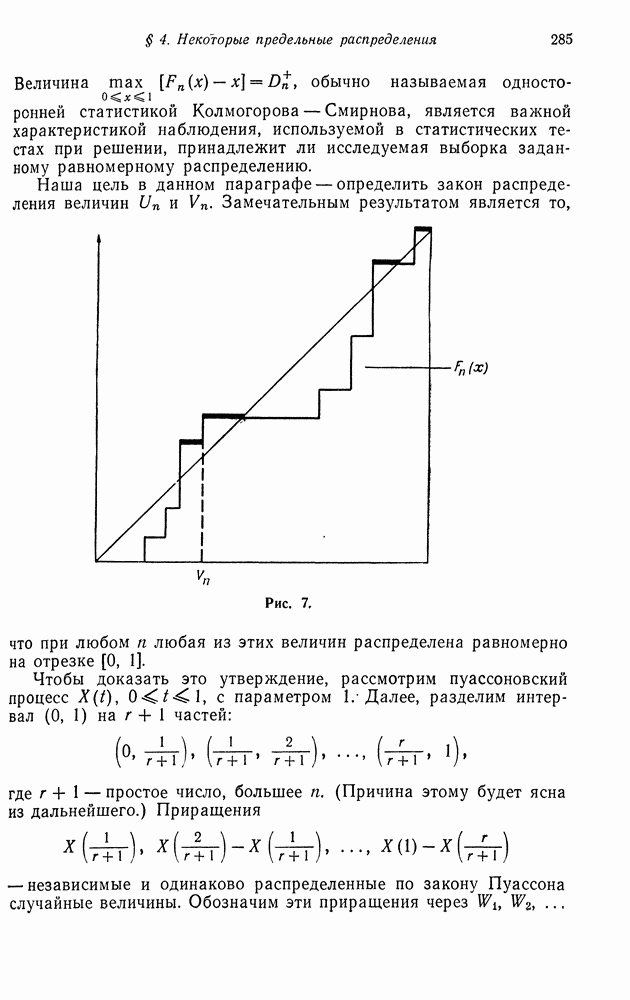
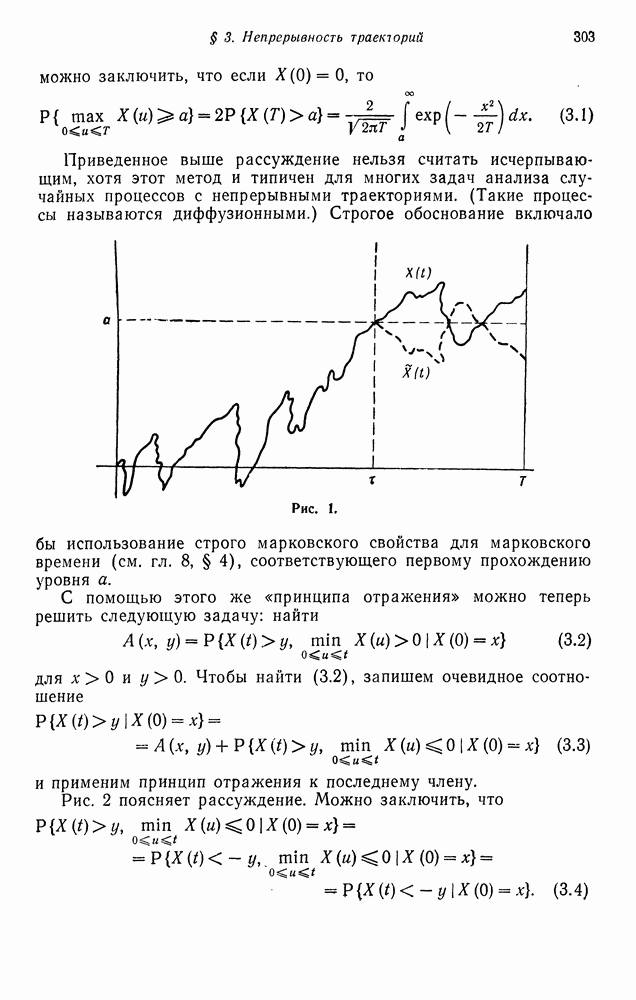
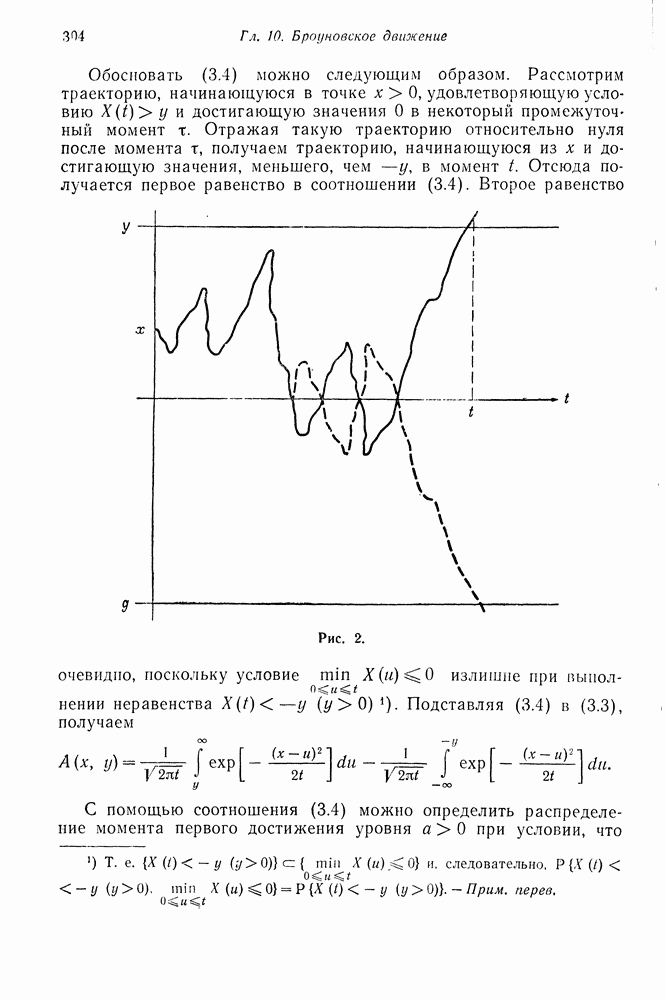
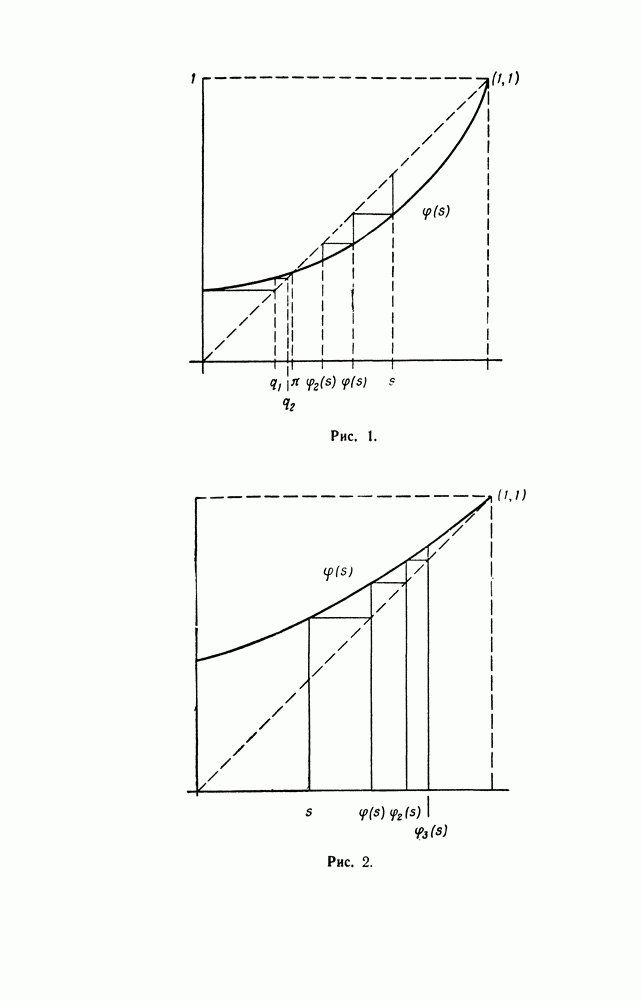
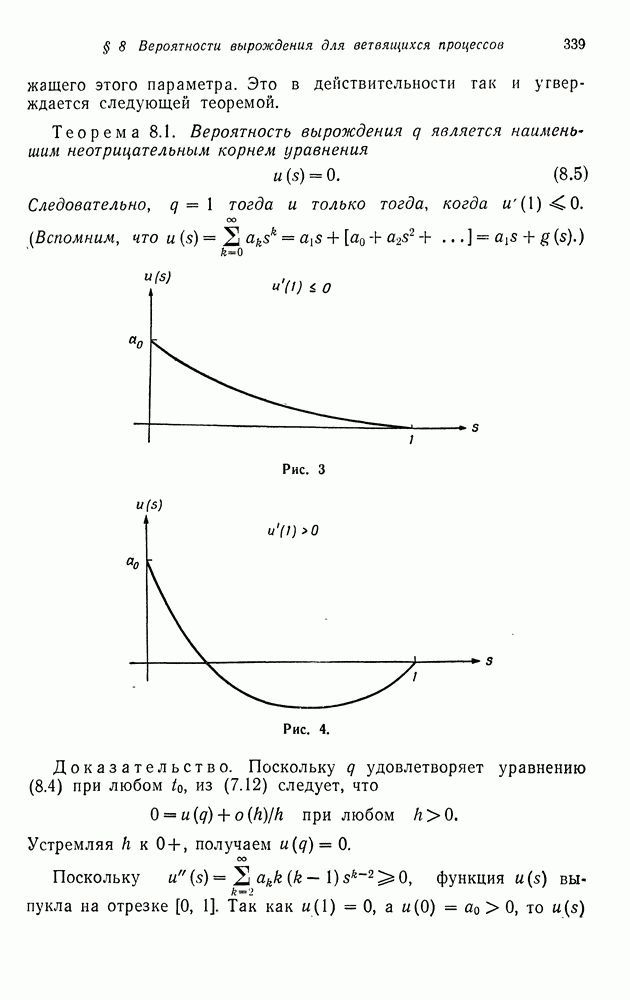
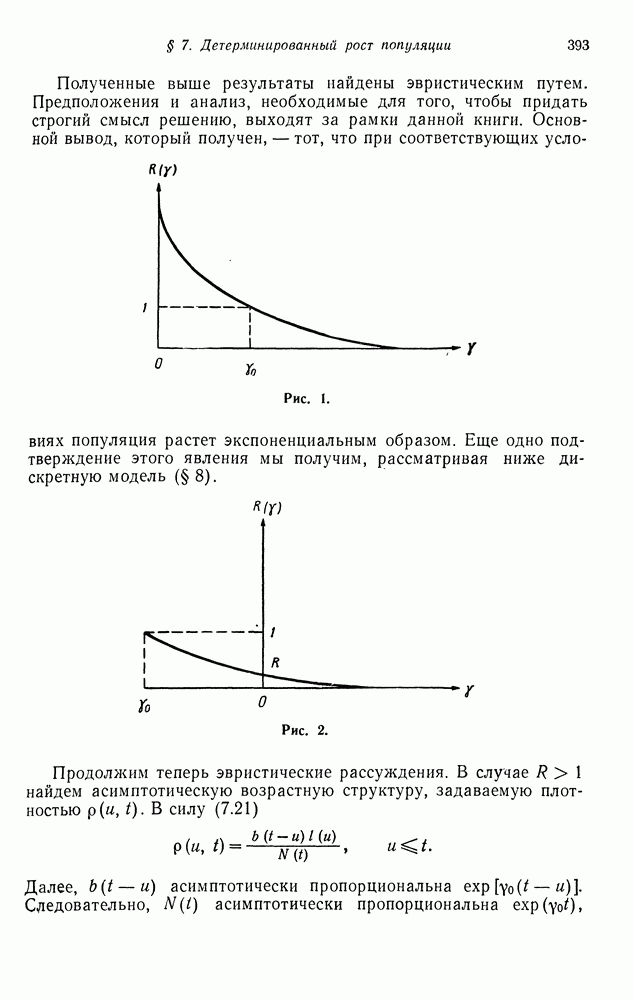
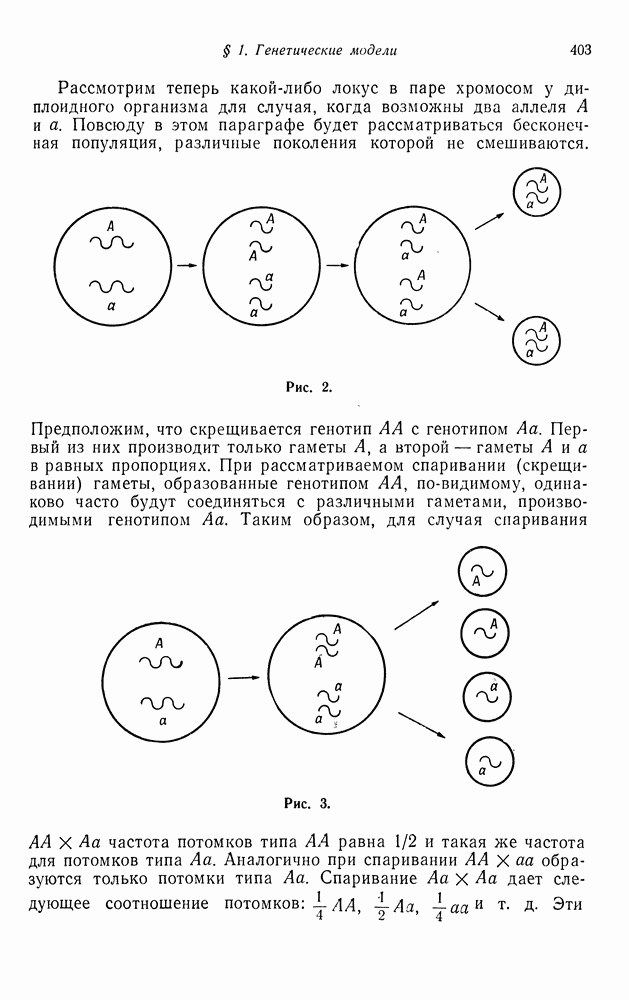
§ 4. МАРКОВСКИЕ ПРОЦЕССЫ, ПОРОЖДАЕМЫЕ ПРЯМЫМ ПРОИЗВЕДЕНИЕМ ВЕТВЯЩИХСЯ ПРОЦЕССОВ
Существует тесная связь между теорией ветвящихся процессов и некоторыми вероятностными моделями частот, используемыми в генетике и экологии популяций. Мы введем в рассмотрение класс конечных цепей Маркова специального вида, который включает в себя как частные случаи цепи, рассмотренные в § 2 гл. 2 в качестве примеров приложения к генетике.
Пусть  Тогда марковский ветвящийся процесс определяется как цепь Маркова, для которой элемент матрицы переходных вероятностей
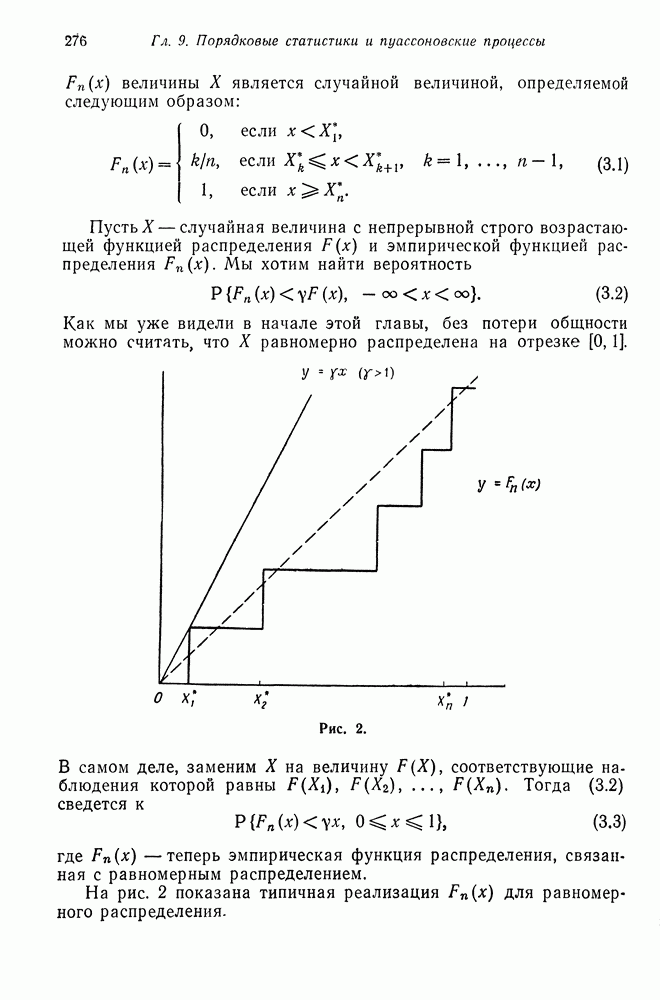
Тогда марковский ветвящийся процесс определяется как цепь Маркова, для которой элемент матрицы переходных вероятностей  определяется, как коэффициент при
определяется, как коэффициент при  в разложении
в разложении  (см. гл. 11). Вероятностное распределение, определяемое последовательностью
(см. гл. 11). Вероятностное распределение, определяемое последовательностью  называется распределением числа потомков для одного индивидуума, а
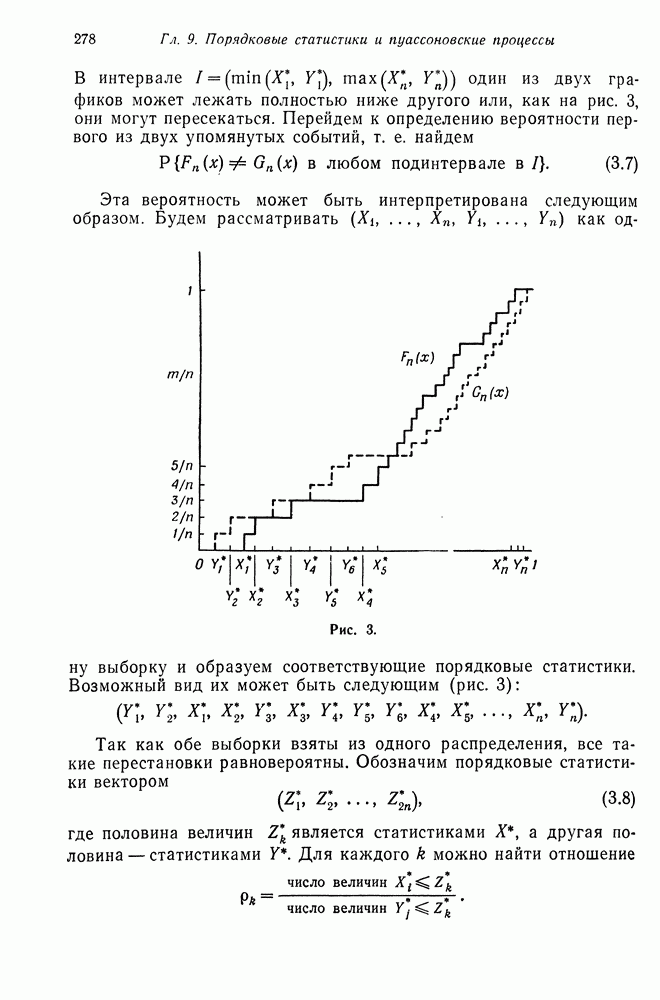
называется распределением числа потомков для одного индивидуума, а  вероятностной производящей функцией для числа независимо порожденных потомков
вероятностной производящей функцией для числа независимо порожденных потомков  индивидуумов. Аналогично процесс
индивидуумов. Аналогично процесс  является ветвящимся, если
является ветвящимся, если  где
где  число индивидуумов в текущем поколении,
число индивидуумов в текущем поколении,  число потомков
число потомков  индивидуума в текущем поколении, а
индивидуума в текущем поколении, а  число индивидуумов в последующем поколении. Случайные величины
число индивидуумов в последующем поколении. Случайные величины  являются независимо распределенными с общей вероятностной производящей функцией
являются независимо распределенными с общей вероятностной производящей функцией 
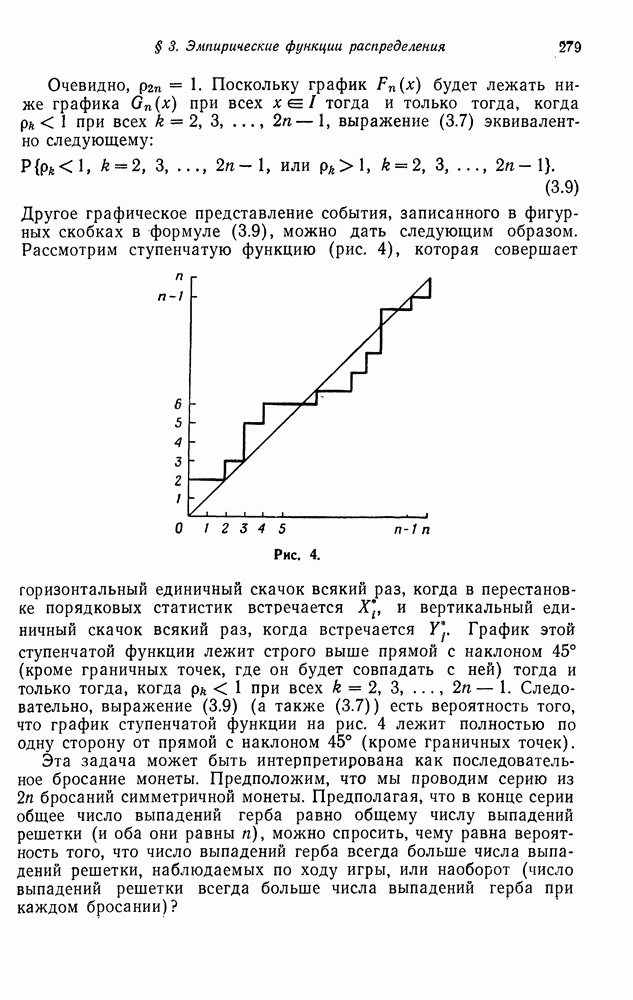
Рассмотрим теперь две популяции типов  каждая из которых независимым образом размножается, как ветвящийся процесс. Это — двумерный процесс, развертывающийся как последовательность пар случайных величин
каждая из которых независимым образом размножается, как ветвящийся процесс. Это — двумерный процесс, развертывающийся как последовательность пар случайных величин  где
где  соответственно числа типов А и а в
соответственно числа типов А и а в  поколении. Состояние процесса описывается парой
поколении. Состояние процесса описывается парой  где
где  текущие значения числа типов А и а соответственно. В такой формулировке компоненты
текущие значения числа типов А и а соответственно. В такой формулировке компоненты  образуют независимые ветвящиеся процессы. Поэтому можно назвать
образуют независимые ветвящиеся процессы. Поэтому можно назвать  прямым произведением ветвящихся процессов
прямым произведением ветвящихся процессов 
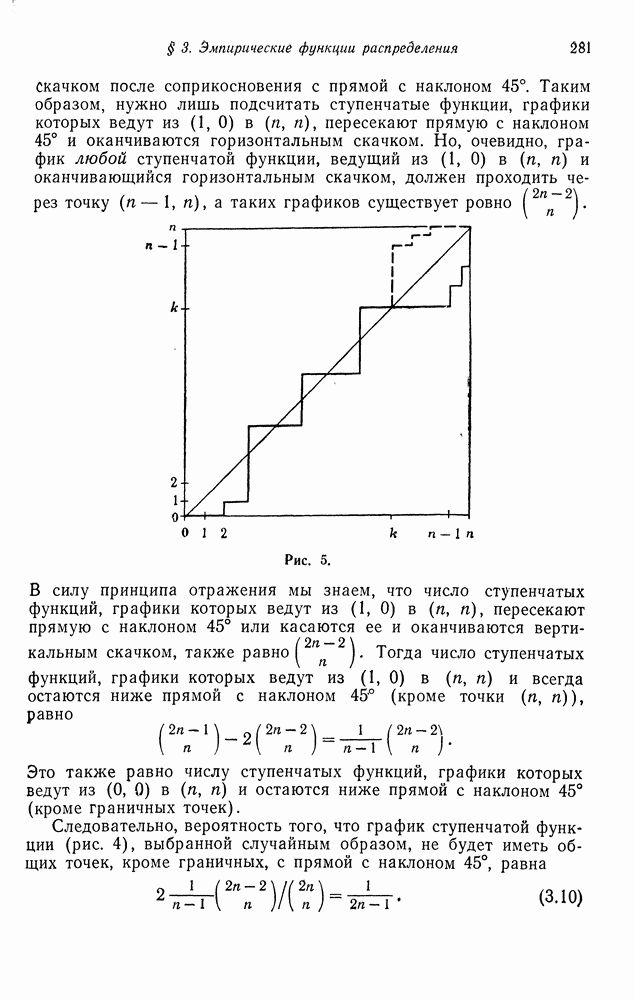
Пусть  производящая функция распределения числа потомков индивидуумов типа
производящая функция распределения числа потомков индивидуумов типа  производящая функция распределения числа потомков индивидуумов типа а. Если индивидуумы развиваются независимо друг от друга, то компоненты матрицы переходных вероятностей вычисляются по формуле
производящая функция распределения числа потомков индивидуумов типа а. Если индивидуумы развиваются независимо друг от друга, то компоненты матрицы переходных вероятностей вычисляются по формуле

Можно сформулировать марковскую модель частот, связанную с ветвящимся процессом для двух типов индивидуумов, в которой размер популяции постоянен во времени. Имея это в виду, найдем сначала вероятность того, что в поколении потомков имеется  индивидуумов типа А при условии, что общее число потомков равно
индивидуумов типа А при условии, что общее число потомков равно  и текущее поколение состоит из
и текущее поколение состоит из  индивидуумов типа
индивидуумов типа  индивидуумов типа а:
индивидуумов типа а:

Здесь использован тот факт, что  производящая функция общего числа потомков, не различаемых по типам. Если
производящая функция общего числа потомков, не различаемых по типам. Если  то можно считать выражение (4.1) определяющим матрицу переходных вероятностей
то можно считать выражение (4.1) определяющим матрицу переходных вероятностей  конечной цепи Маркова с пространством состояний
конечной цепи Маркова с пространством состояний 
Отношение номера состояния к числу  можно интерпретировать как частоту типа А во всей популяции. Построенная цепь Маркова, следовательно, является дискретной вероятностной моделью, которая описывает изменения частот типов в популяции постоянного размера, состоящей из двух типов индивидуумов. Изменения частоты в каждом из поколений определяются видом матрицы переходных вероятностей цепи Маркова.
можно интерпретировать как частоту типа А во всей популяции. Построенная цепь Маркова, следовательно, является дискретной вероятностной моделью, которая описывает изменения частот типов в популяции постоянного размера, состоящей из двух типов индивидуумов. Изменения частоты в каждом из поколений определяются видом матрицы переходных вероятностей цепи Маркова.
Размер популяции  является параметром, который часто для упрощения обозначений будет опускаться. Таким образом,
является параметром, который часто для упрощения обозначений будет опускаться. Таким образом,

Марковскую цепь, характеризуемую матрицей переходных вероятностей (4.2), иногда называют цепью Маркова, порожденной прямым произведением марковских ветвящихся процессов.
Мы проиллюстрируем возможности данного класса цепей Маркова на трех простейших примерах и укажем их место в прикладных задачах.
(1) В качестве первого примера рассмотрим прямое произведение пуассоновских ветвящихся процессов, когда

При этом

Если  то получаем выражение
то получаем выражение

не зависящее от  . Это в точности совпадает с вероятностной моделью биномиального выбора (пример
. Это в точности совпадает с вероятностной моделью биномиального выбора (пример  § 2 гл. 2), предложенной С. Райтом для изучения флуктуаций частоты гена в гаплоидной популяции при воздействии случайного дрейфа. Если
§ 2 гл. 2), предложенной С. Райтом для изучения флуктуаций частоты гена в гаплоидной популяции при воздействии случайного дрейфа. Если  то (4.3) сводится к модели отбора Райта, в которой
то (4.3) сводится к модели отбора Райта, в которой  коэффициент отбора, выражающий преимущество индивидуумов типа А над индивидуумами типа а.
коэффициент отбора, выражающий преимущество индивидуумов типа А над индивидуумами типа а.
В общем случае, соответствующем формуле (4.1), мы скажем, что отбор имеет место, если  т. е. если вероятностные производящие функции числа потомков двух типов различны.
т. е. если вероятностные производящие функции числа потомков двух типов различны.
(2) Пусть 
Порожденная цепь Маркова (4.2) в данном случае принимает вид

Интерпретация этой цепи Маркова как модели полисомного на следования дана в примере 3 § 2 гл. 2.
(3) Пусгь

Тогда, используя разложение

мы видим, что порожденная цепь Маркова имеет следующие переходные вероятности

Эта цепь Маркова может интерпретироваться как модель роста гетерозиготной популяции (см. стр. 321).
Явления миграции и мутации могут быть учтены в частотной модели, представляемой порожденной марковской цепью, следующим образом. Сначала рассмотрим явление иммиграции. Предположим, что кроме размножения происходит процесс иммиграции в систему независимо от текущего размера популяции. Пусть  вероятностные производящие функции числа иммигрантов типов А и а, поступающих в систему за время одного поколения соответственно. Совместная производящая функция
вероятностные производящие функции числа иммигрантов типов А и а, поступающих в систему за время одного поколения соответственно. Совместная производящая функция  Для вероятностей числа индивидуумов типов А и а в популяции в следующем поколении равна
Для вероятностей числа индивидуумов типов А и а в популяции в следующем поколении равна  Здесь
Здесь

Для того чтобы построить порожденную цепь Маркова, введем условие  Порожденная марковская цепь имеет переходные вероятности
Порожденная марковская цепь имеет переходные вероятности

Явление мутации можно учесть в порожденной цепи Маркова двумя способами. Предположим, что каждый индивидуум типа А может мутировать в индивидуум тина а с вероятностью  и каждый индивидуум типа а может мутировать в индивидуум типа А с вероятностью
и каждый индивидуум типа а может мутировать в индивидуум типа А с вероятностью  По отношению к мутации индивидуумы являются независимыми. Для того чтобы образовать следующее поколение, мы можем постулировать, что процесс мутации следует за процессом роста (ветвящегося размножения) или наоборот. Эти два постулата приводят к различным цепям Маркова.
По отношению к мутации индивидуумы являются независимыми. Для того чтобы образовать следующее поколение, мы можем постулировать, что процесс мутации следует за процессом роста (ветвящегося размножения) или наоборот. Эти два постулата приводят к различным цепям Маркова.
Наиболее просто описать функционирование модели мутации непосредственно в терминах соответствующих вероятностных про изводящих функций. Рассмотрим сначала случай, когда мутация происходит после воспроизведения. Пусть  как и прежде, — производящие функции числа потомков одного родителя типов А и а соответственно. Предположим, что при следующем процессе воспроизведения каждый потомок индивидуума типа А (типа а) может производить индивидуумы обоих видов с производящей функцией
как и прежде, — производящие функции числа потомков одного родителя типов А и а соответственно. Предположим, что при следующем процессе воспроизведения каждый потомок индивидуума типа А (типа а) может производить индивидуумы обоих видов с производящей функцией  Другими словами, мы постулируем две фазы процесса воспроизведения: первая соответствует обычному процессу размножения, в котором потомки являются копиями родителей, в то время как вторая фаза соответствует своего рода процессу трансформации и роста, когда один индивидуум одного типа может превращаться в индивидуумов другого типа, т. е. коэффициент
Другими словами, мы постулируем две фазы процесса воспроизведения: первая соответствует обычному процессу размножения, в котором потомки являются копиями родителей, в то время как вторая фаза соответствует своего рода процессу трансформации и роста, когда один индивидуум одного типа может превращаться в индивидуумов другого типа, т. е. коэффициент  в разложении
в разложении  является вероятностью того, что на второй фазе потомок типа А произведет
является вероятностью того, что на второй фазе потомок типа А произведет  индивидуумов типа
индивидуумов типа  индивидуумов типа а. Производящая функция, описывающая вид популяции потомков одного индивидуума типа
индивидуумов типа а. Производящая функция, описывающая вид популяции потомков одного индивидуума типа  (типа а) после обеих фаз воспроизводства, равна
(типа а) после обеих фаз воспроизводства, равна 
Механизм мутации получается при следующем ограничении:

При этом вторая фаза воспроизводства сводится к процессу трансформации С вероятностью  потомок типа А сохраняет свой тип, а с вероятностью
потомок типа А сохраняет свой тип, а с вероятностью  мутирует в тип а. Аналогичную интерпретацию имеет и производящая функция
мутирует в тип а. Аналогичную интерпретацию имеет и производящая функция 
В результате производящая функция популяции потомков, полученных при размножении одного индивидуума типа А и подверженных затем мутационному давлению, равна

Аналогичным образом можно описать популяцию потомков одного родителя типа а, учитывая мутационное давление, с помощью производящей функции

С другой стороны, если мутационные превращения происходят до воспроизводства, производящие функции принимают вид

Проведенное выше обсуждение приводит к следующей более общей конструкции.
Мы постулируем, что каждый индивидуум типа А может производить потомков обоих типов. Обозначим вероятностную производящую функцию размера популяции потомков через  Аналогично предположим, что индивидуум типа а может производить индивидуумов обоих типов, и пусть
Аналогично предположим, что индивидуум типа а может производить индивидуумов обоих типов, и пусть  — производящая функция популяции его потомков. Пусть
— производящая функция популяции его потомков. Пусть  производящая функция числа индивидуумов типов А и а, иммигрирующих в систему в течение каждого периода. Пусть
производящая функция числа индивидуумов типов А и а, иммигрирующих в систему в течение каждого периода. Пусть  результирующий двумерный ветвящийся процесс. Вероятностная производящая функция величины
результирующий двумерный ветвящийся процесс. Вероятностная производящая функция величины  при начальных условиях
при начальных условиях  равна
равна

Матрица переходных вероятностей порожденной цепи Маркова при условии, что размер популяции фиксирован числом  вычисляется обычным образом. Получим
вычисляется обычным образом. Получим

В частности, если

то переходные вероятности (4.12) порожденной цепи Маркова равны

Эта конечная цепь Маркова была предложена С. Райтом для изучения флуктуаций частоты гена при учете отбора, мутации, миграции и генетического отбора.