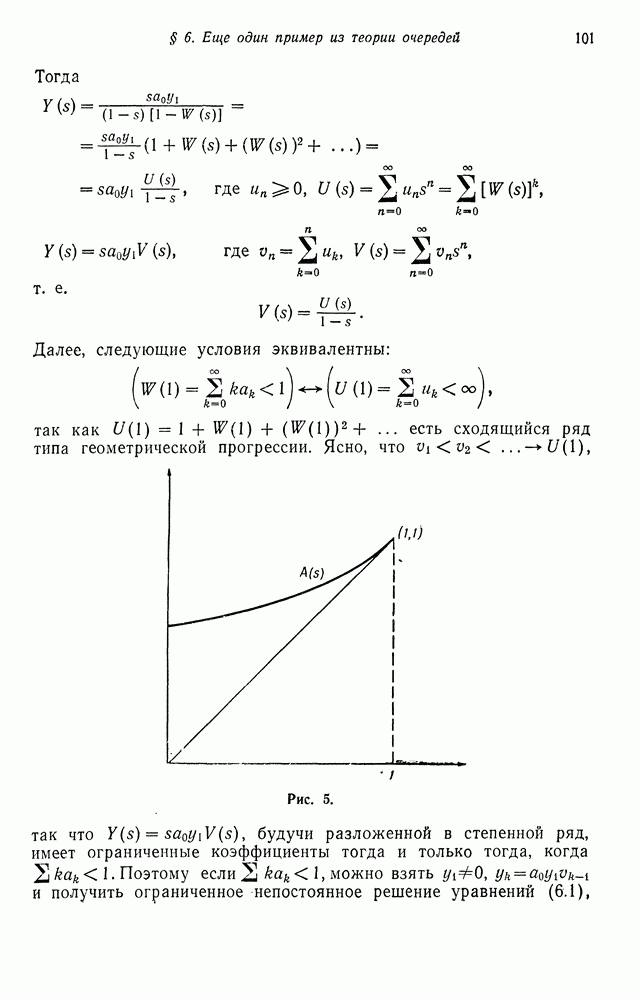
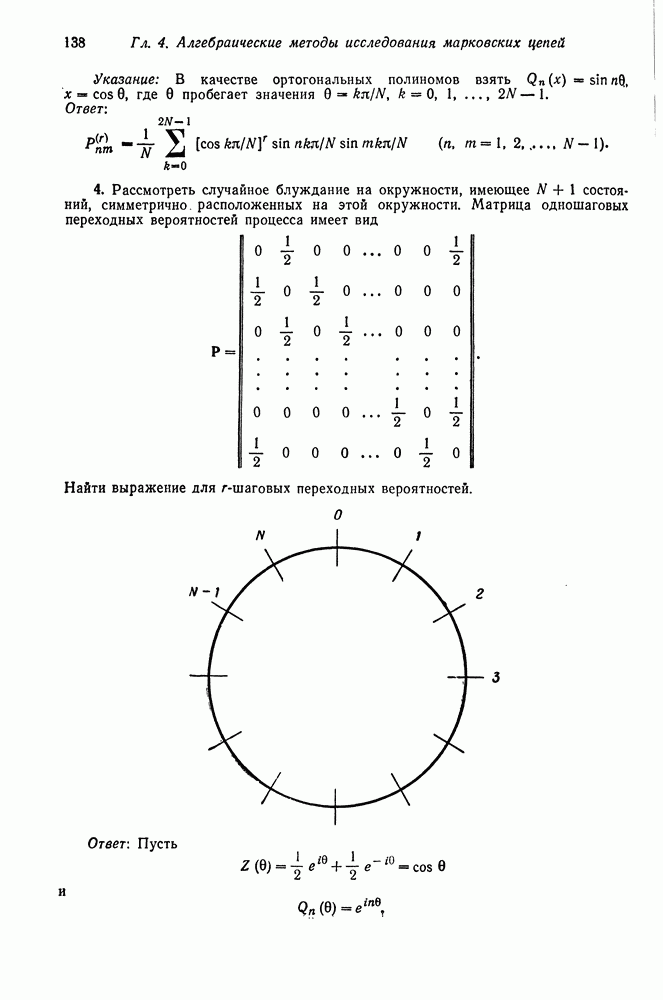
Б. Спаривание сибсов
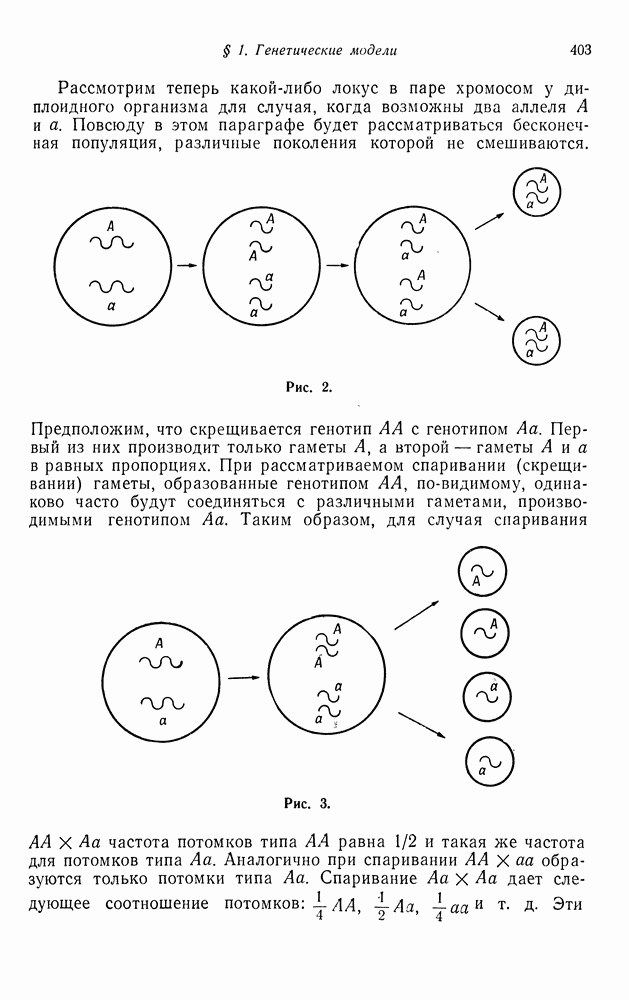
В следующей модели рассматривается спаривание потомков одних и тех же родителей, или спаривание сибсов. Опять целью является определение предельных частот различных генотипов, участвующих в спаривании. Однако вместо непосредственного рассмотрения частот генотипов мы проследим за флуктуациями частот спаривающихся особей в последовательных поколениях.
Если ограничиться рассмотрением двух аллелей А и а, то существуют три возможных генотипа для каждого родителя и шесть
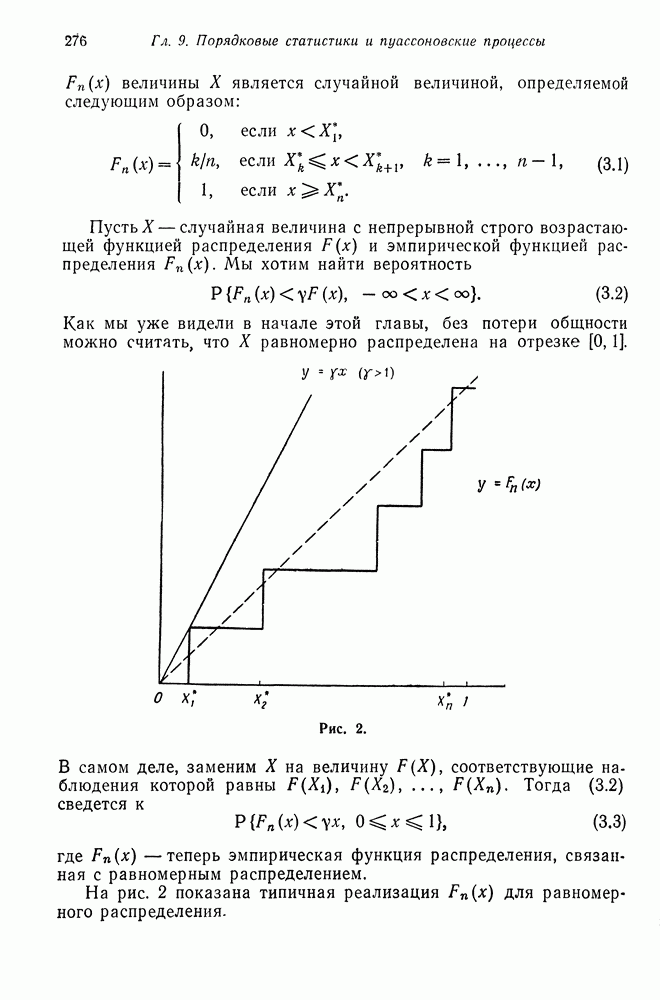
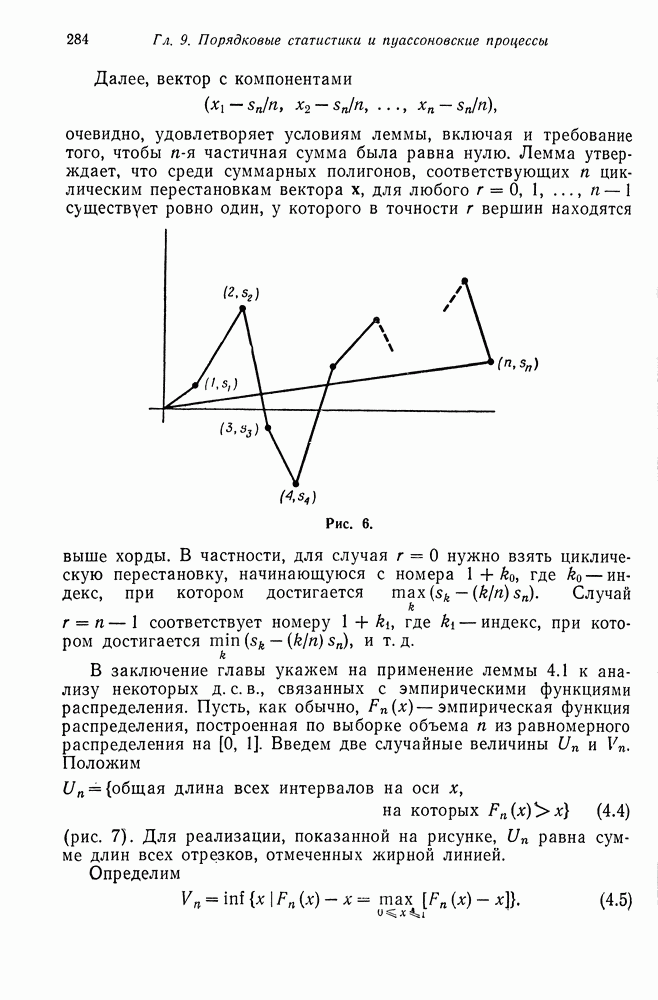
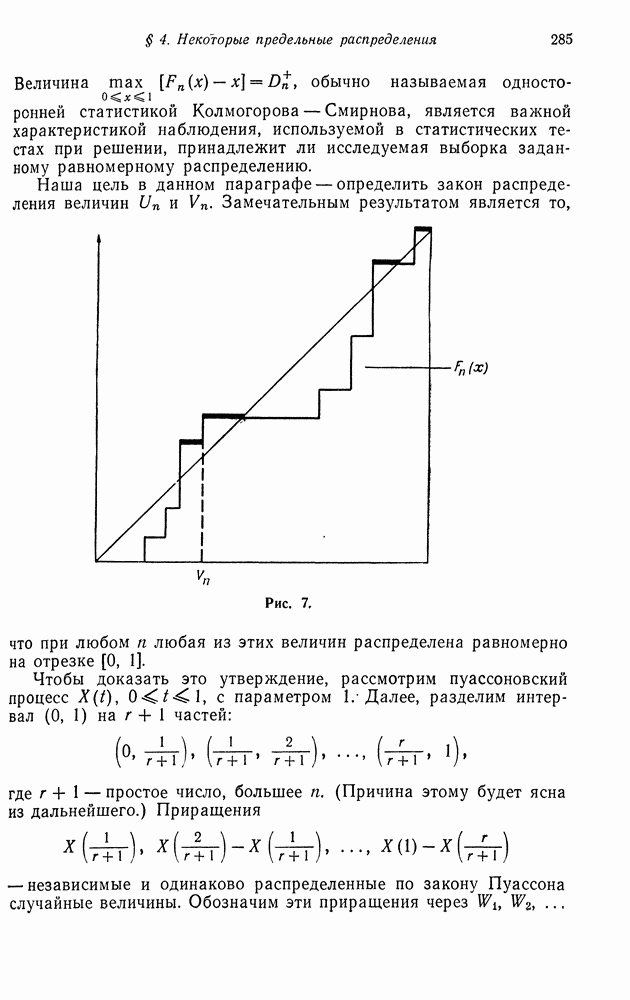
возможных вариантов сочетания, или скрещивания генотипов, т. е. существует шесть различных вариантов спаривания:  Предположим, что частоты каждого из этих вариантов известны. Мы хотим вычислить эти частоты для следующего поколения. Для того чтобы сделать это, мы сейчас найдем распределение генотипов потомков, а затем, предполагая, что спариваться могут только сибсы, определим частоту получаемых при этом различных видов спаривания потомков.
Предположим, что частоты каждого из этих вариантов известны. Мы хотим вычислить эти частоты для следующего поколения. Для того чтобы сделать это, мы сейчас найдем распределение генотипов потомков, а затем, предполагая, что спариваться могут только сибсы, определим частоту получаемых при этом различных видов спаривания потомков.
Например, все дети, получаемые при спаривании  имеют тип
имеют тип  и поэтому среди сибсов допустимым является лишь спаривание вида
и поэтому среди сибсов допустимым является лишь спаривание вида  При спаривании
При спаривании  производятся потомки в отношении —
производятся потомки в отношении —  Частоты различных вариантов спаривания среди сибсов, очевидно, равны
Частоты различных вариантов спаривания среди сибсов, очевидно, равны 
Аналогичным образом можно рассмотреть и другие возможности. Так, при спаривании  потомки производятся в отношении
потомки производятся в отношении  Частоты различных вариантов спаривания среди сибсов можно легко вычислить. Они равны
Частоты различных вариантов спаривания среди сибсов можно легко вычислить. Они равны

Другие сочетания исследуются аналогичным образом.
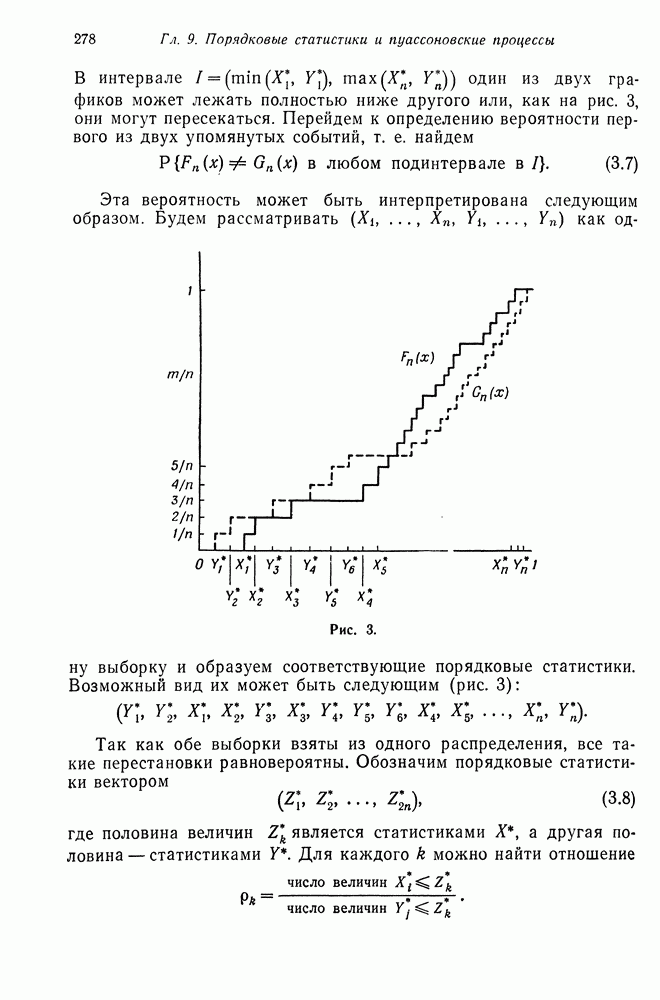
Можно проанализировать изменения частот спаривающихся  от поколения к поколению с помощью цепей Маркова. Для этого рассмотрим цепь Маркова с шестью состояниями, соответствующими шести вариантам спаривания:
от поколения к поколению с помощью цепей Маркова. Для этого рассмотрим цепь Маркова с шестью состояниями, соответствующими шести вариантам спаривания:

Эта цепь Маркова имеет следующую матрицу перехода:

В строках матрицы записаны частоты различных вариантов спаривания среди сибсов, родившихся в результате соответствующего номеру строки варианта спаривания. Имеются два поглощающих состояния: 1 и 6.
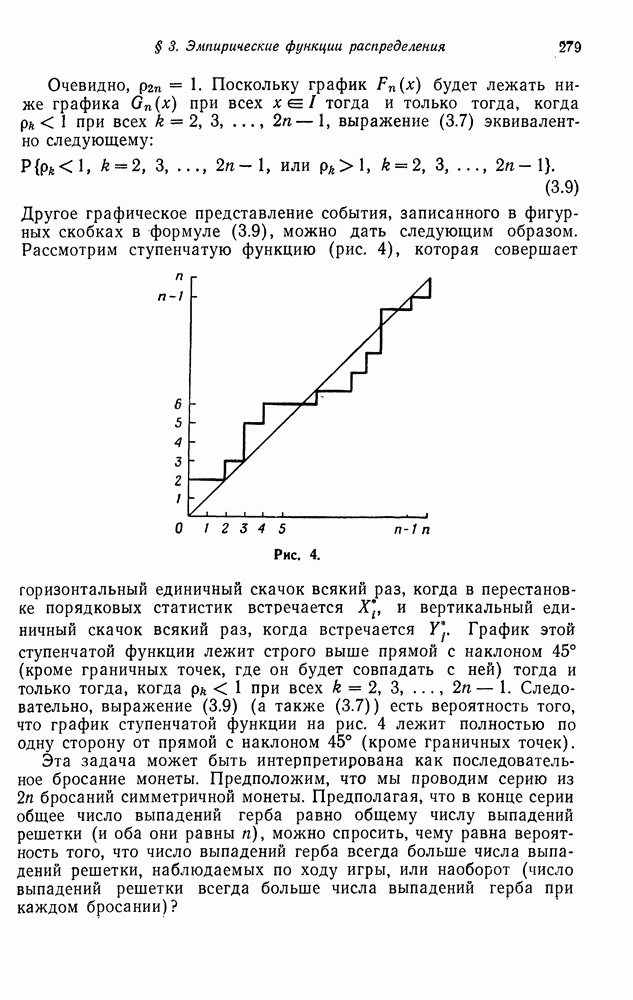
Из теории марковских цепей известно, что в конце концов произойдет поглощение в одном из этих двух состояний. Вероятность поглощения из переходных состояний (состояний 2—5) представляет интерес, поскольку она, очевидно, соответствует той интенсивности, с которой исчезают гетерозиготные генотипы. Эта интенсивность равна, конечно, наибольшему собственному значению матрицы

которая получится при исключении первого и шестого состояний. Поскольку последствия вариантов спаривания  симметричны, можно должным образом объединить первую и четвертую строки матрицы В. При этом получим матрицу
симметричны, можно должным образом объединить первую и четвертую строки матрицы В. При этом получим матрицу

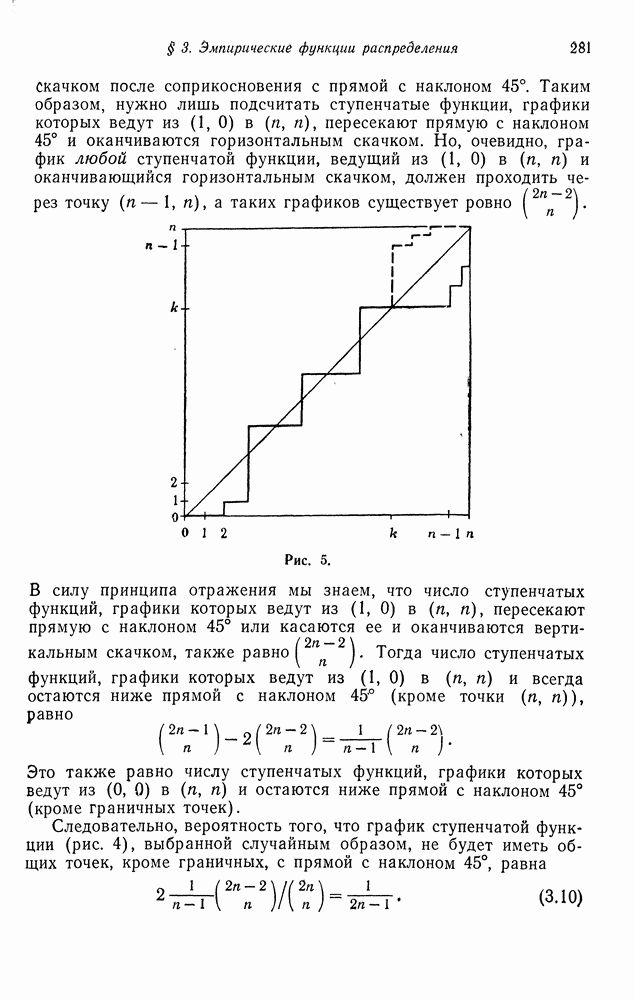
Собственные значения С являются корнями уравнения

а именно 

Следовательно, скорость приближения к гомозиготному состоянию (фиксации) равна

т. е. с каждым поколением частота вариантов спаривания различных генотипов уменьшается в  раз.
раз.
Теперь не представляет труда вычислить при любых начальных частотах  вариантов спаривания финальную частоту
вариантов спаривания финальную частоту
поглощения, или фиксации в состояниях  или
или  Точно так же мы определим частоту фиксации двух гомозиготных генотипов.
Точно так же мы определим частоту фиксации двух гомозиготных генотипов.
Вероятность  фиксации в состоянии 1 (спаривание
фиксации в состоянии 1 (спаривание  при начальном состоянии
при начальном состоянии  является решением уравнения
является решением уравнения

где полагается  Нетрудно найти, что
Нетрудно найти, что

Если начальные частоты различных состояний равны  , то вероятность фиксации в состоянии 1, очевидно, равна
, то вероятность фиксации в состоянии 1, очевидно, равна