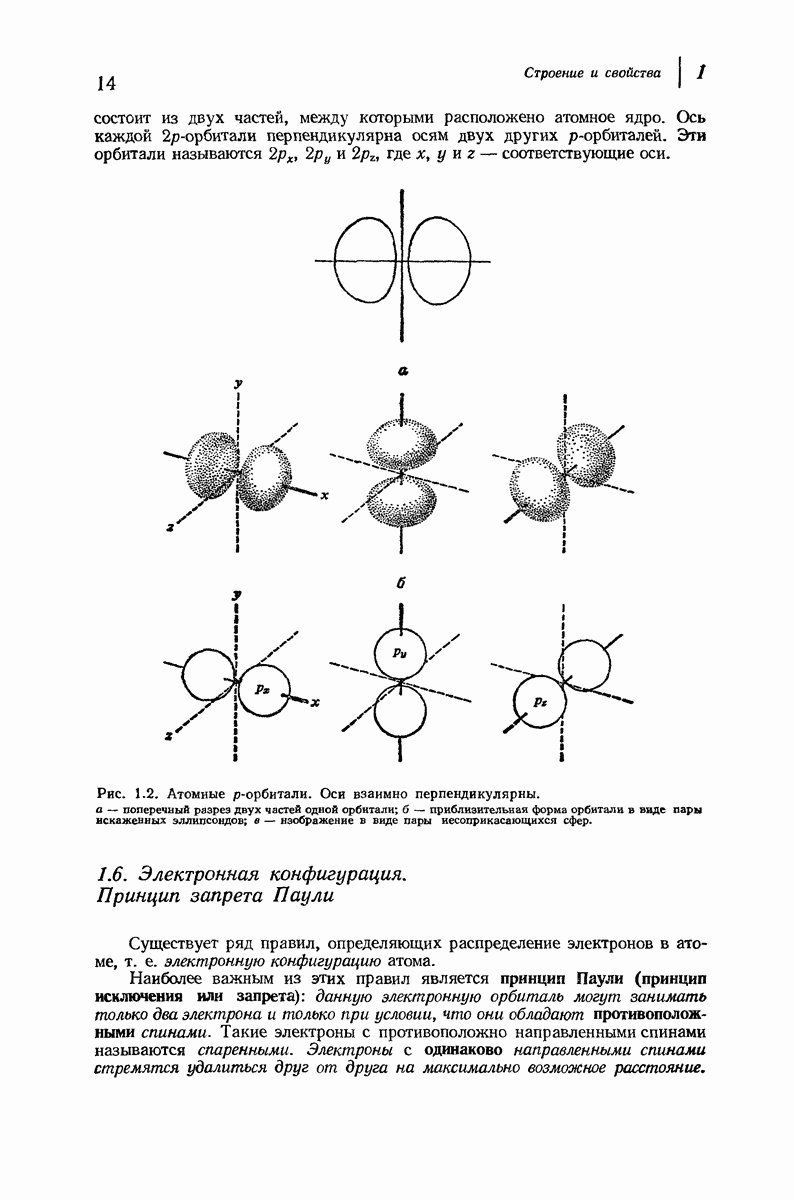
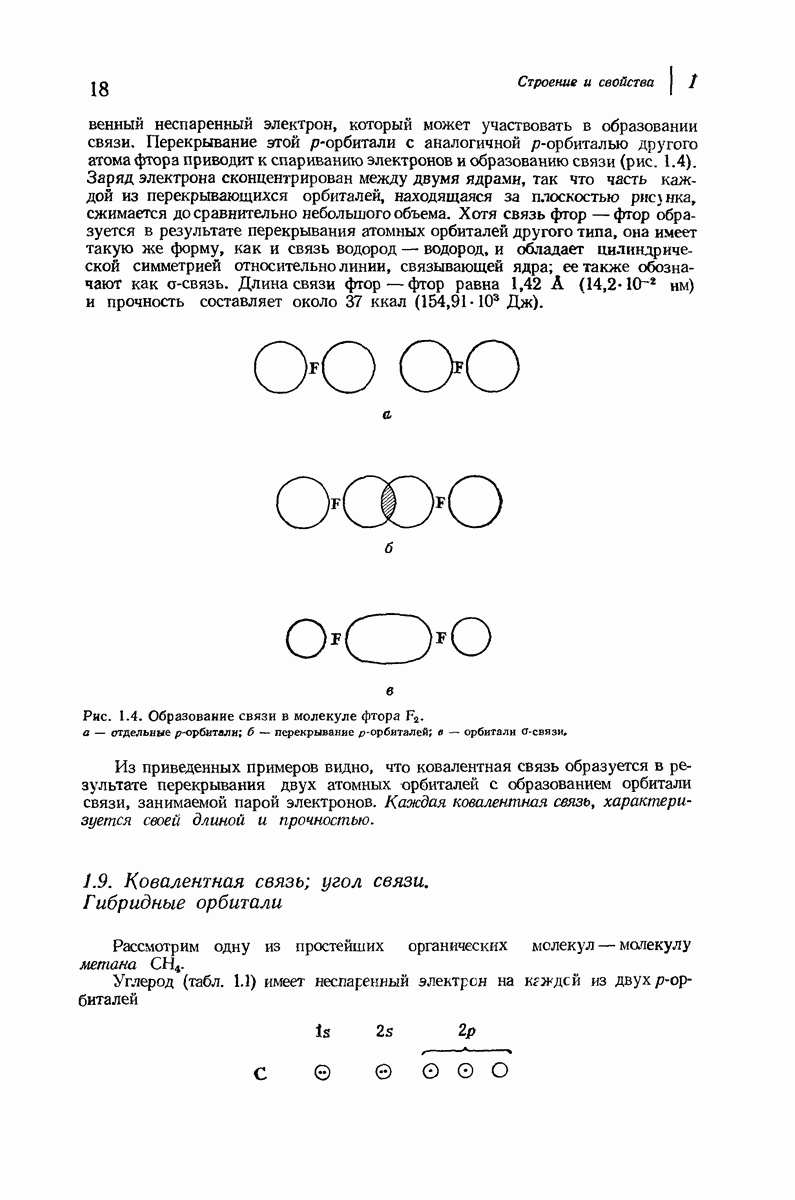
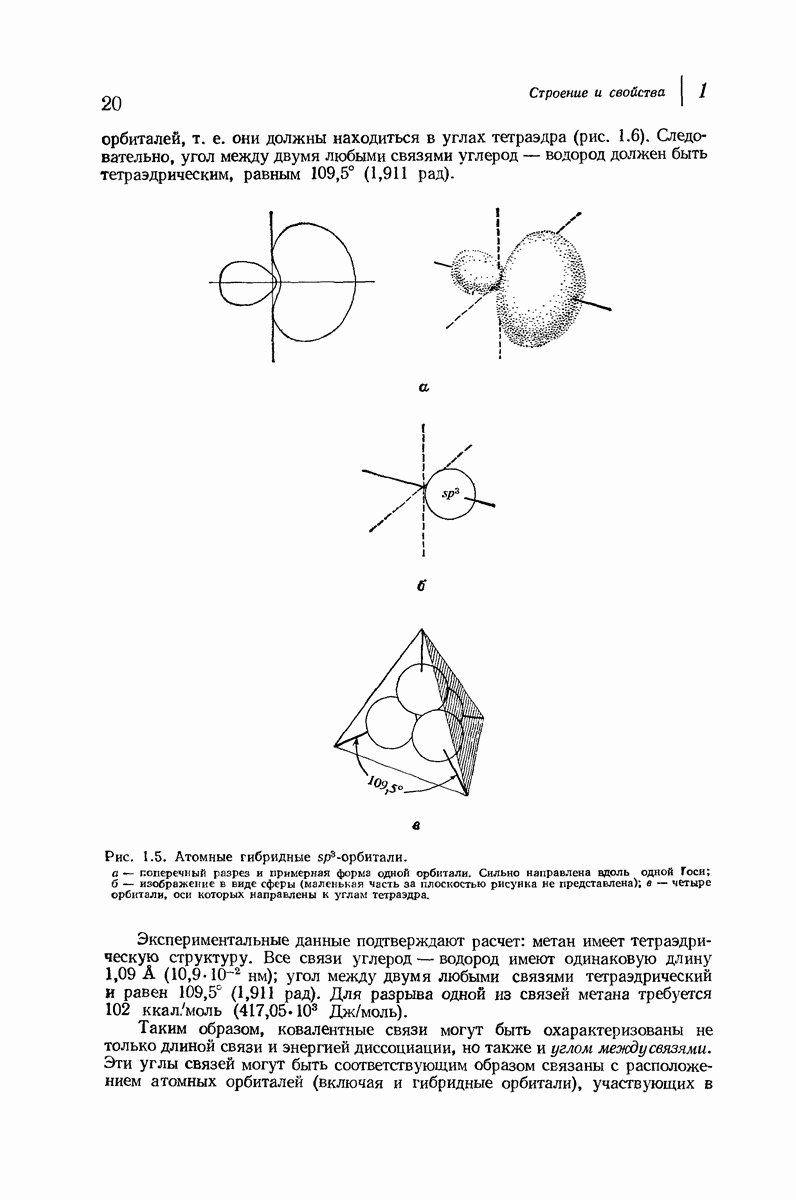
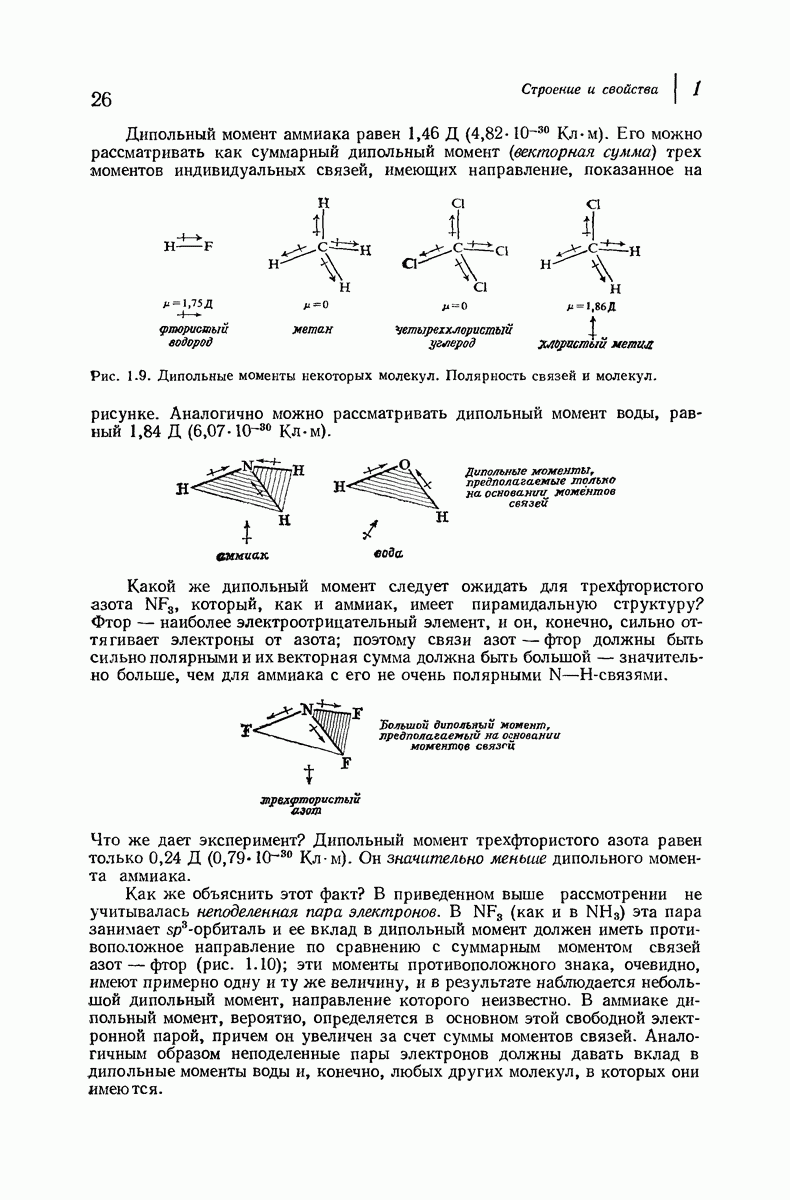
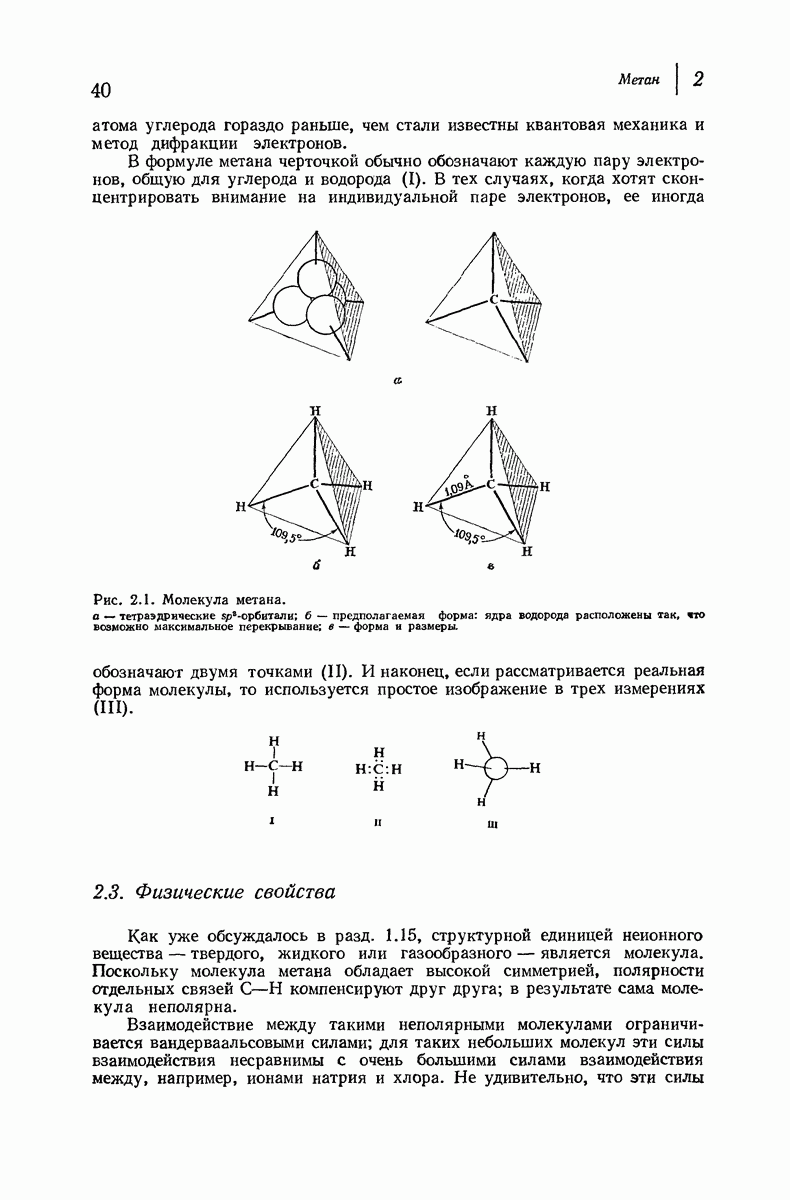
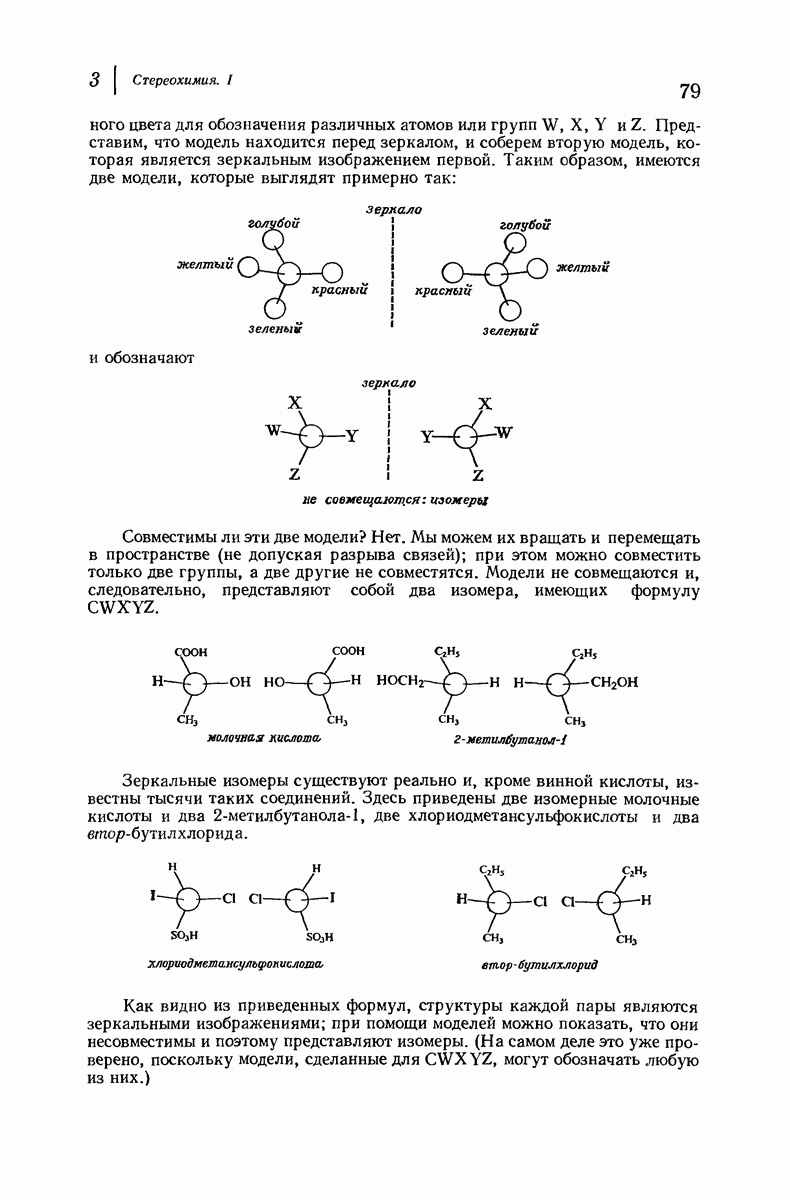
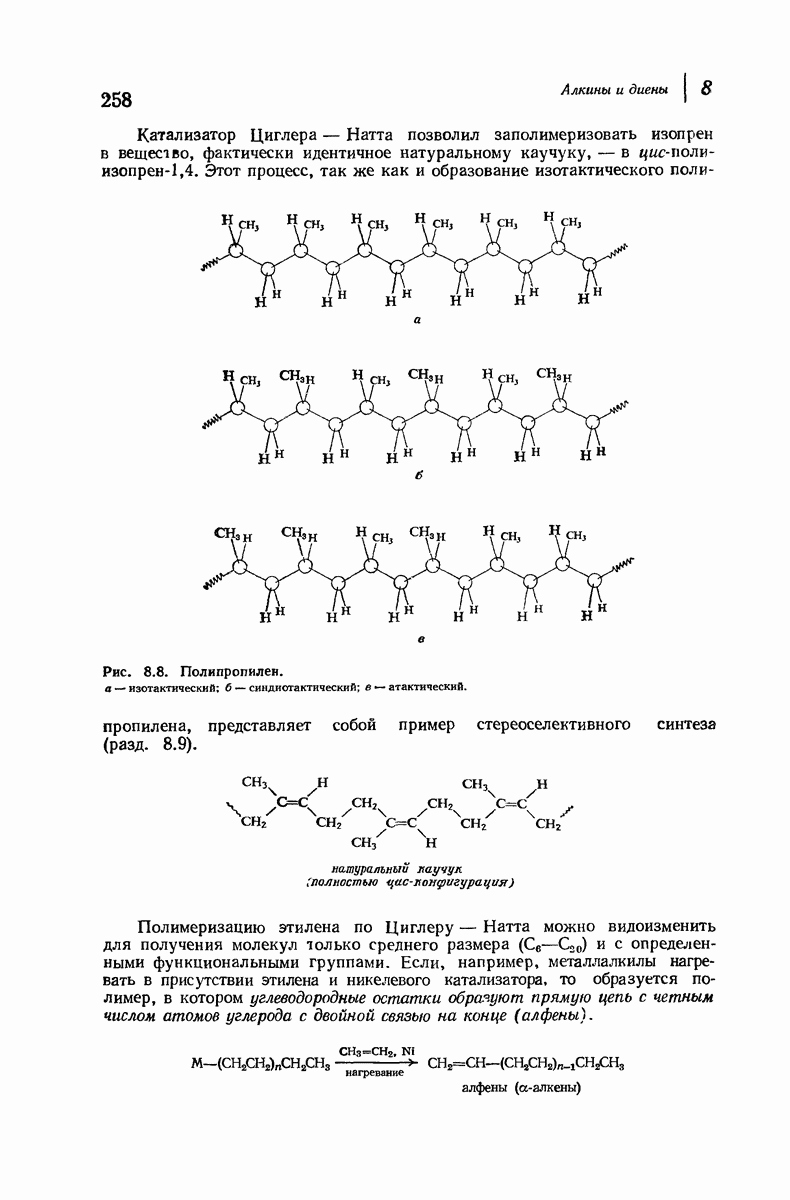
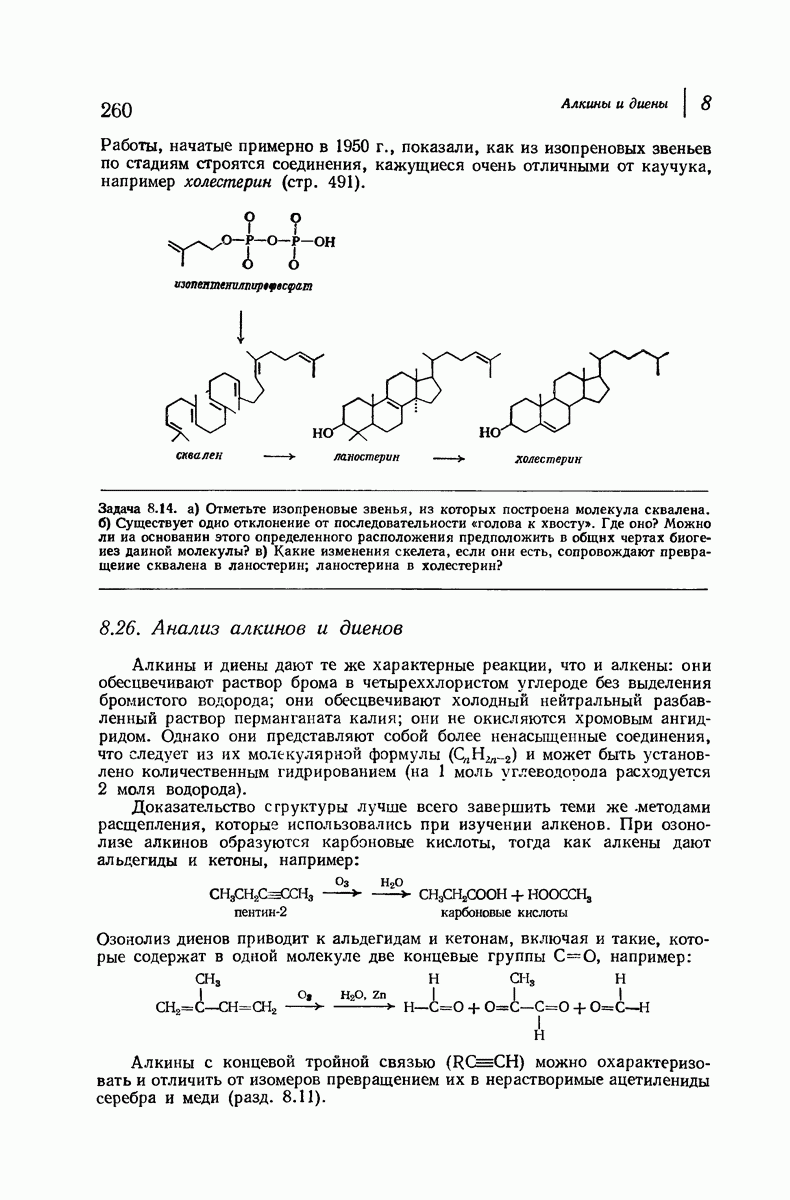
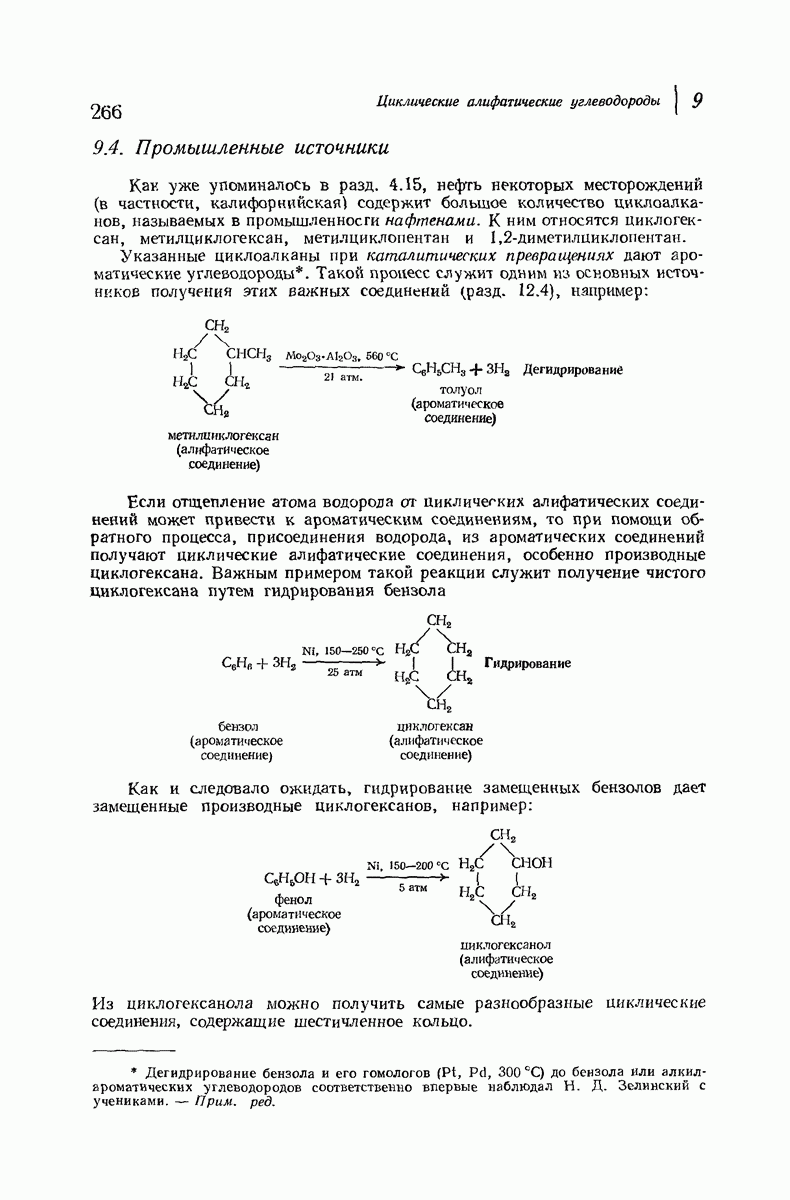
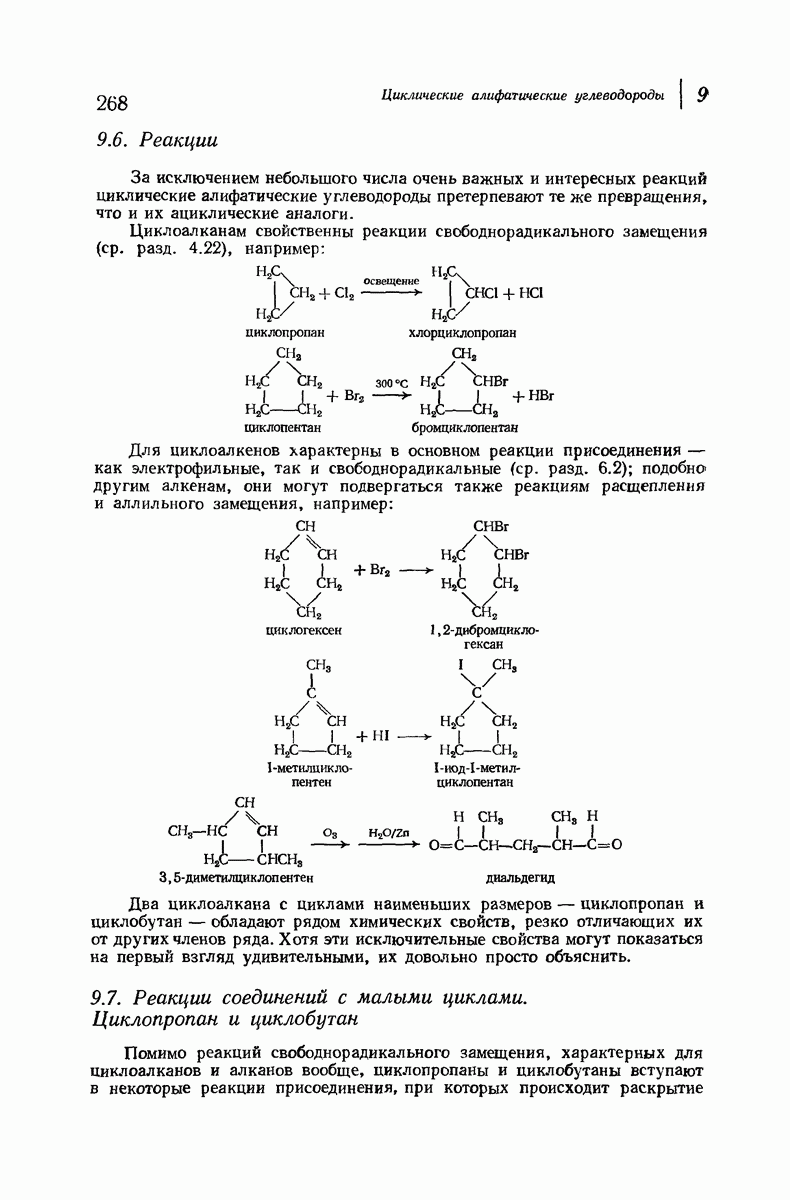
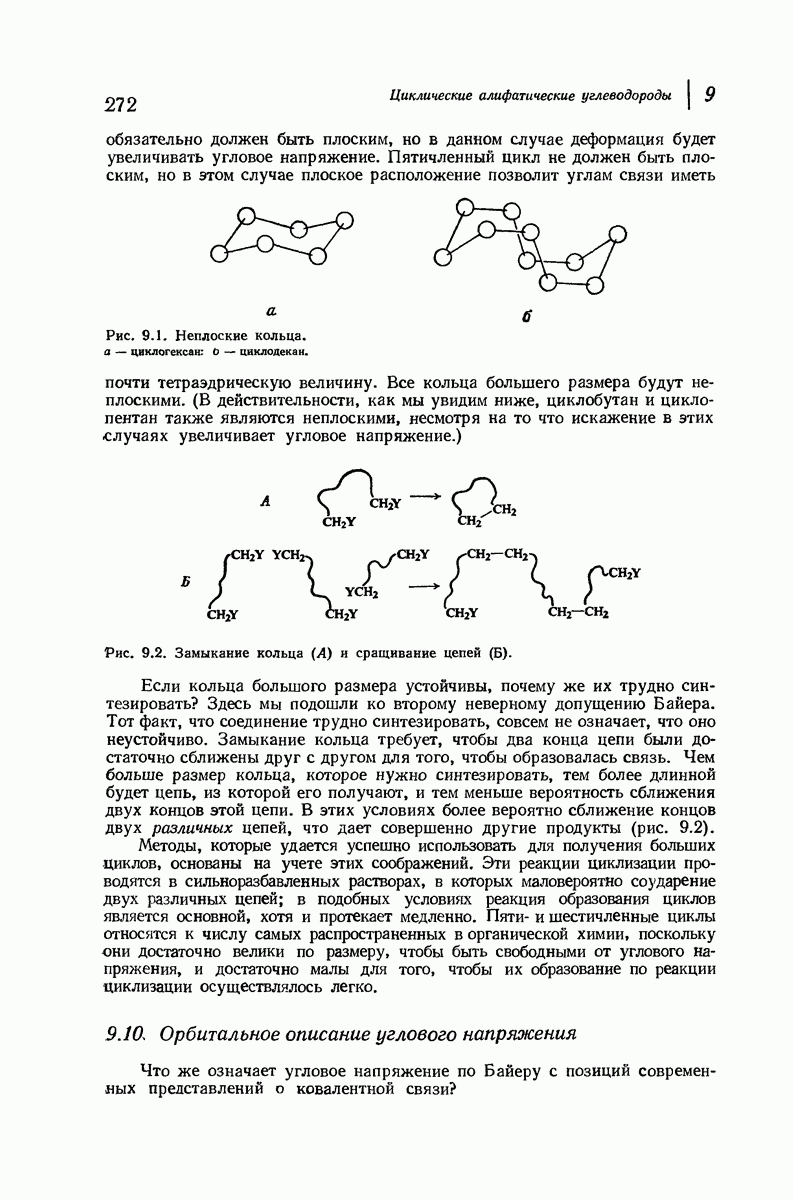
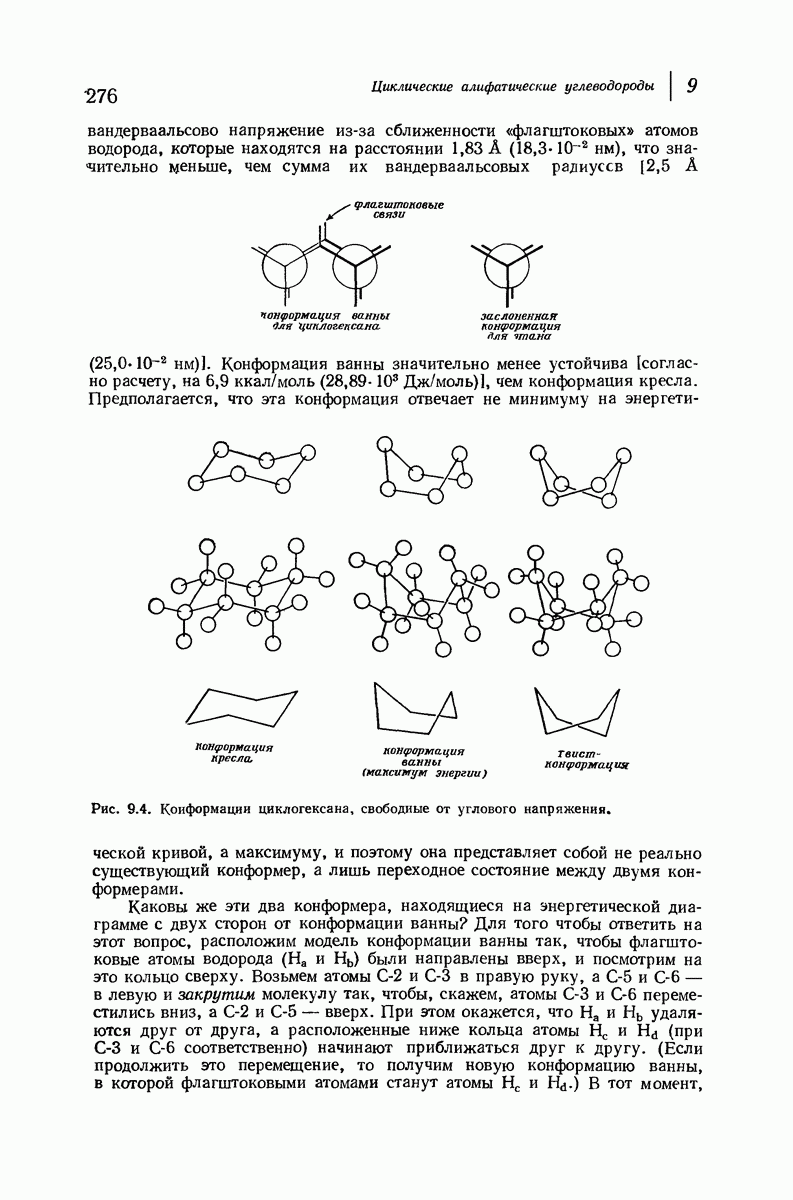
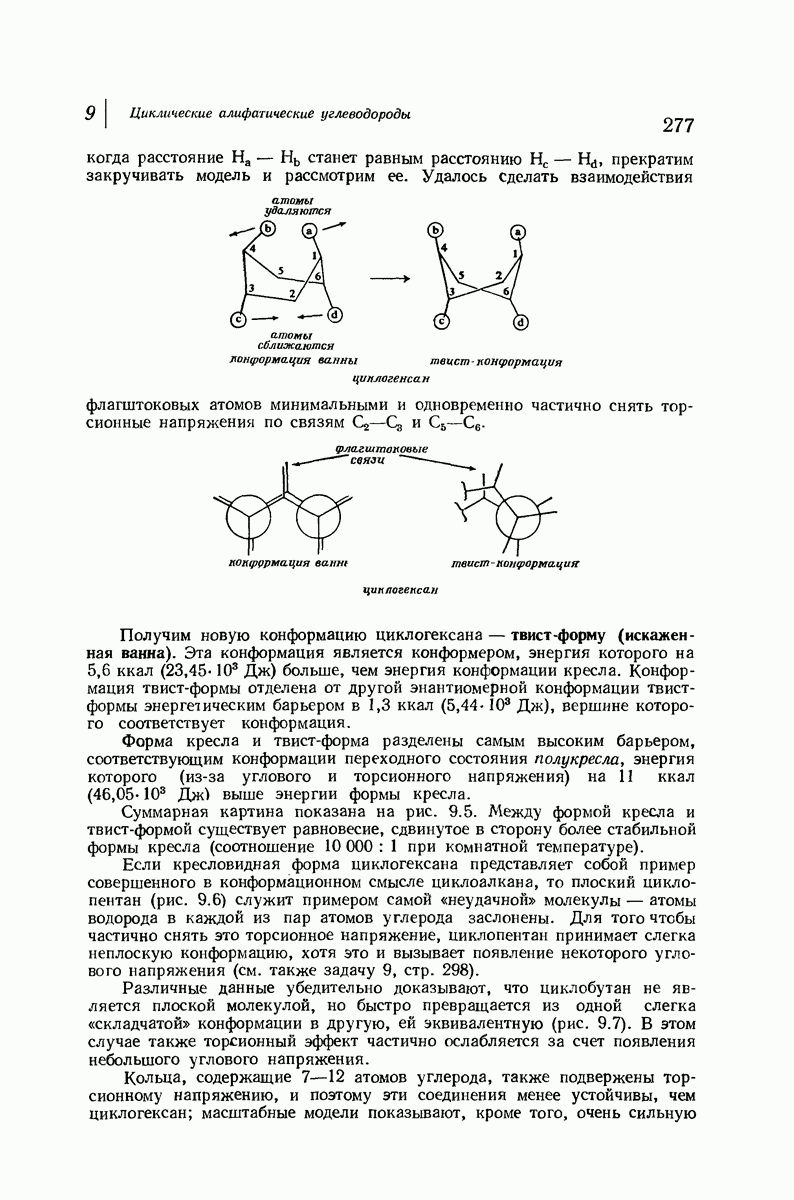
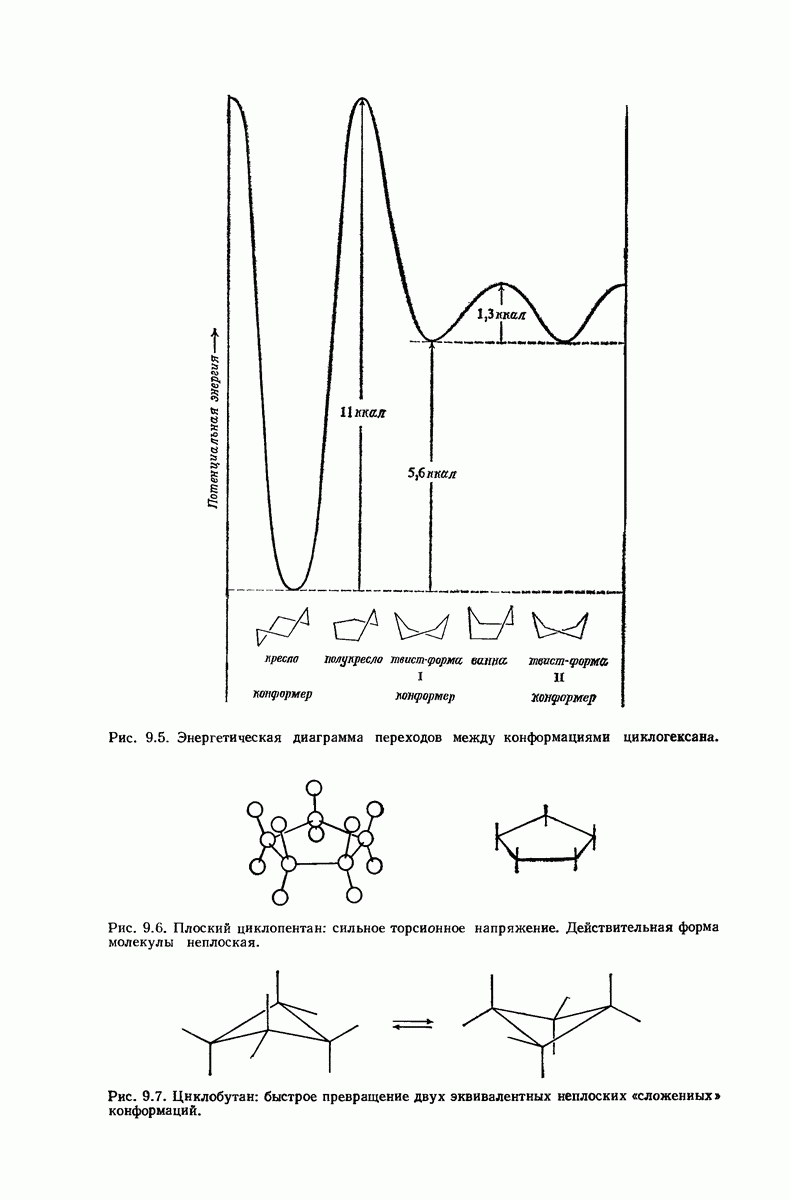
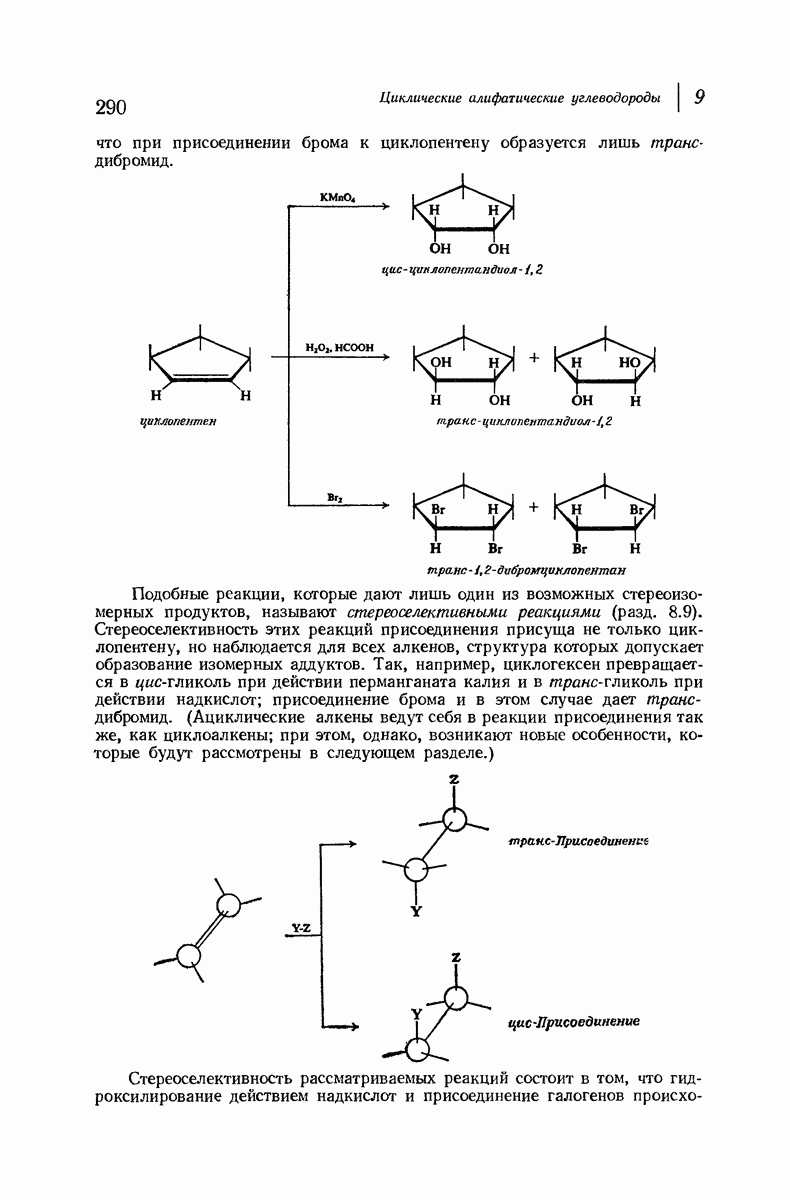
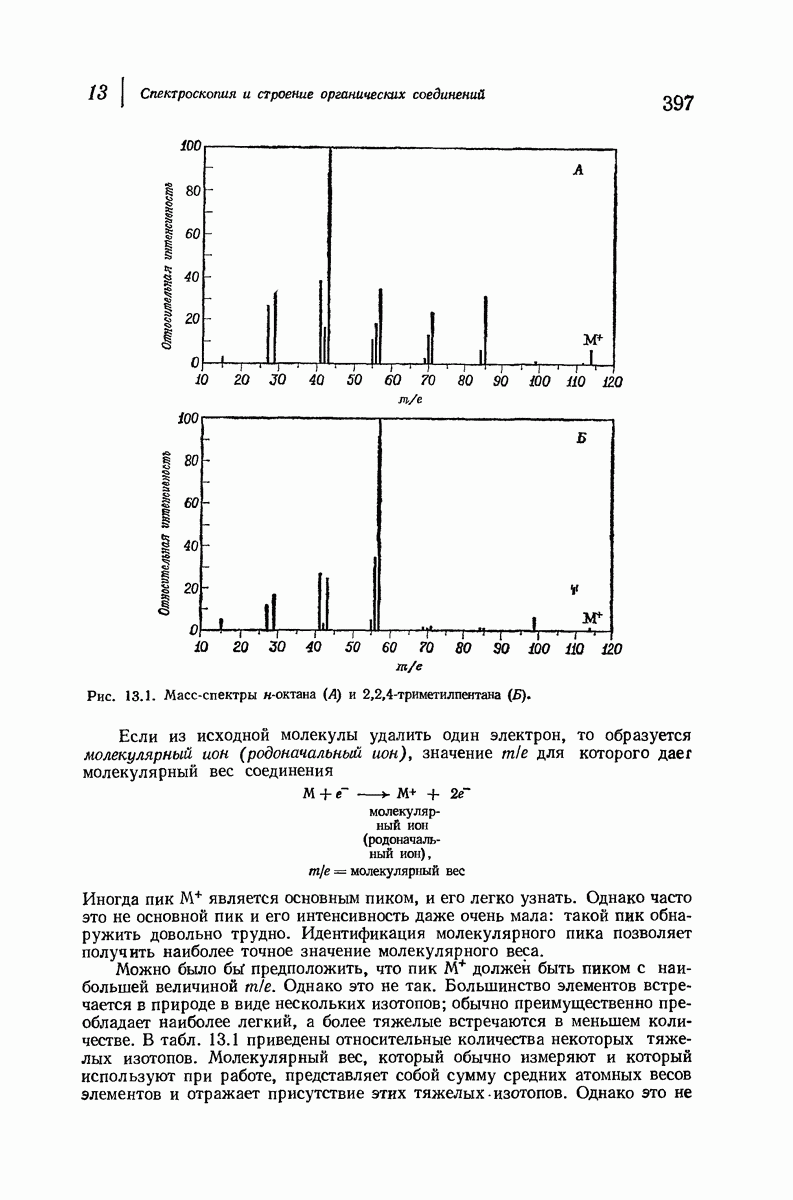
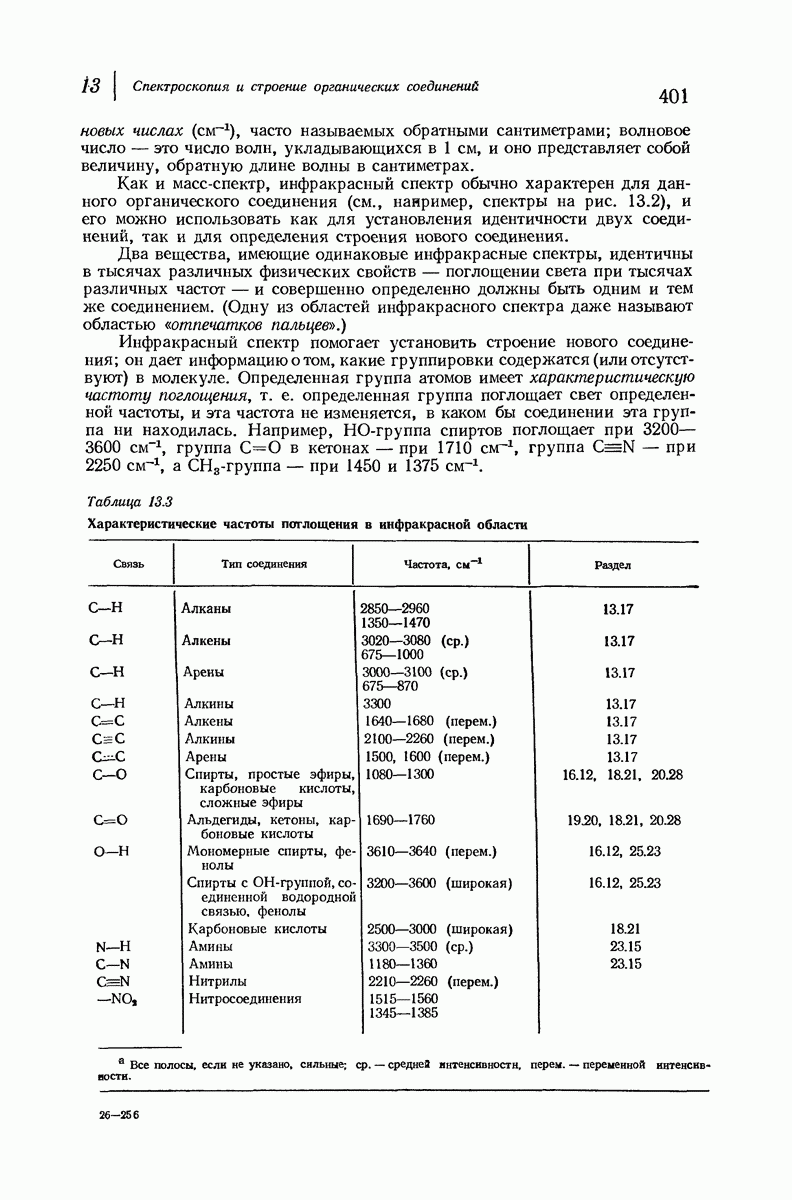
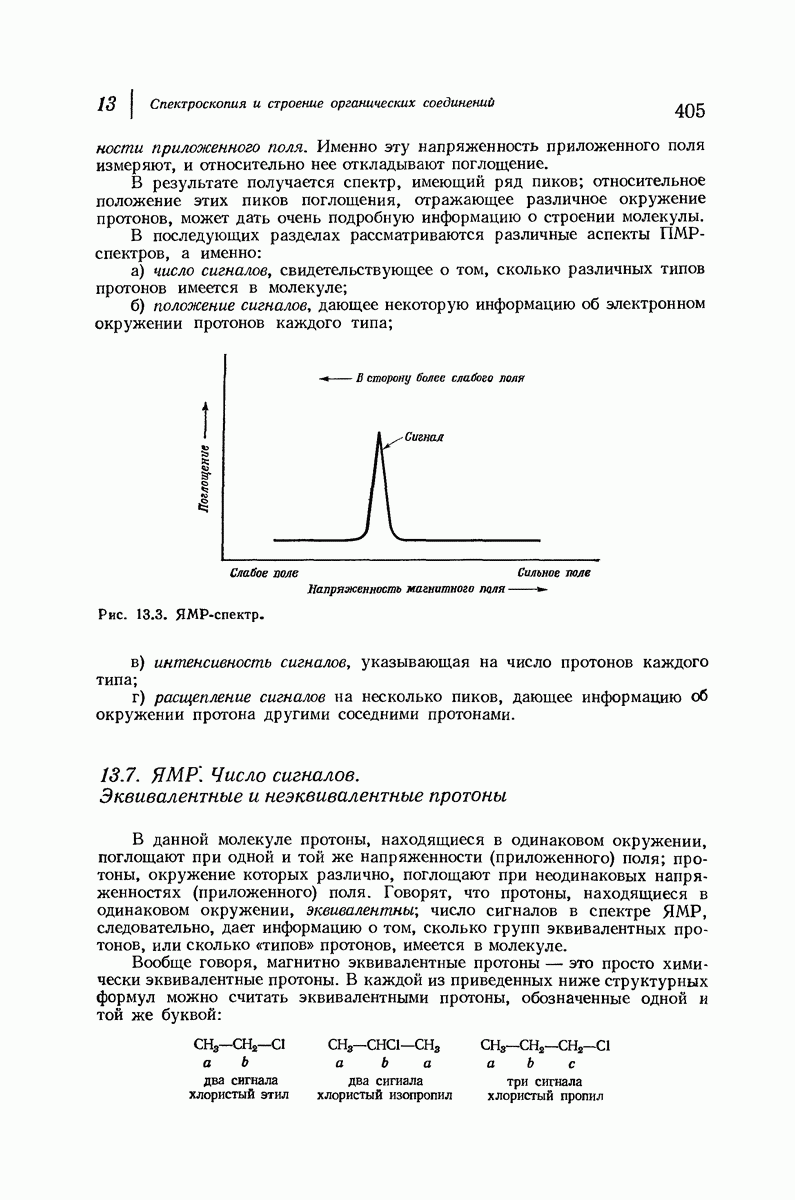
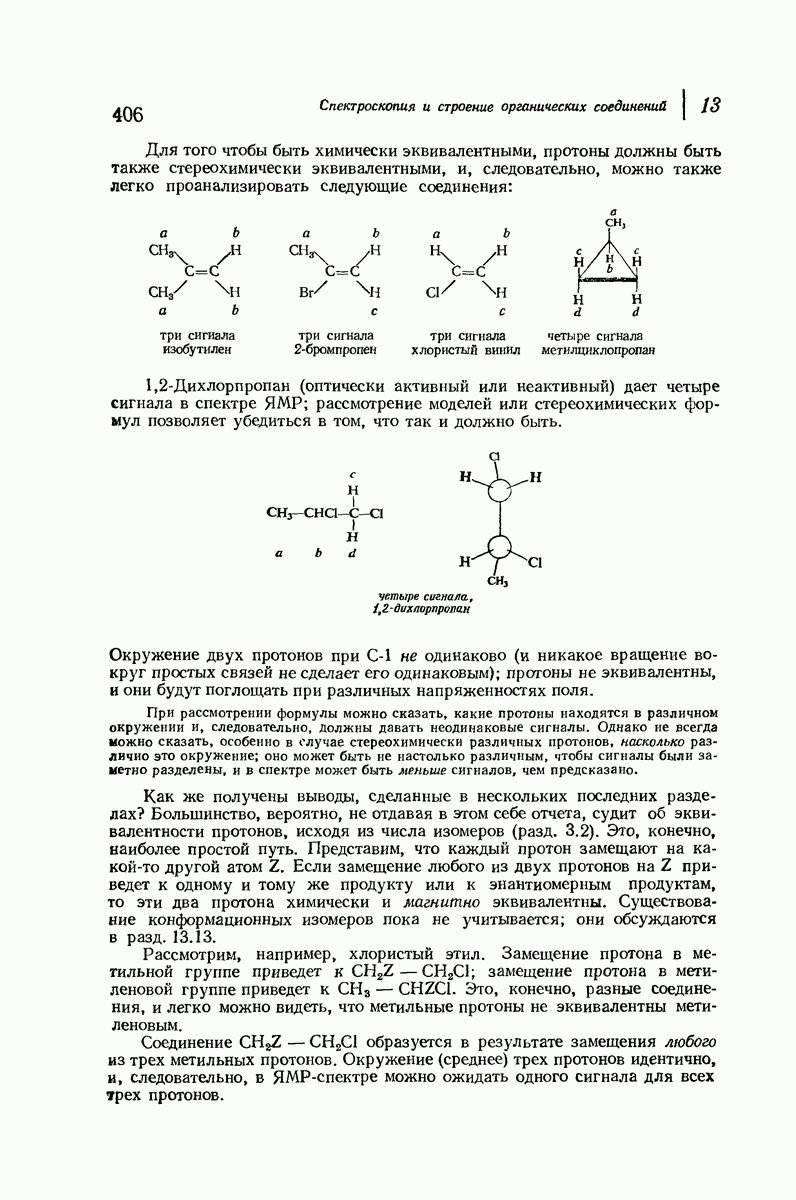
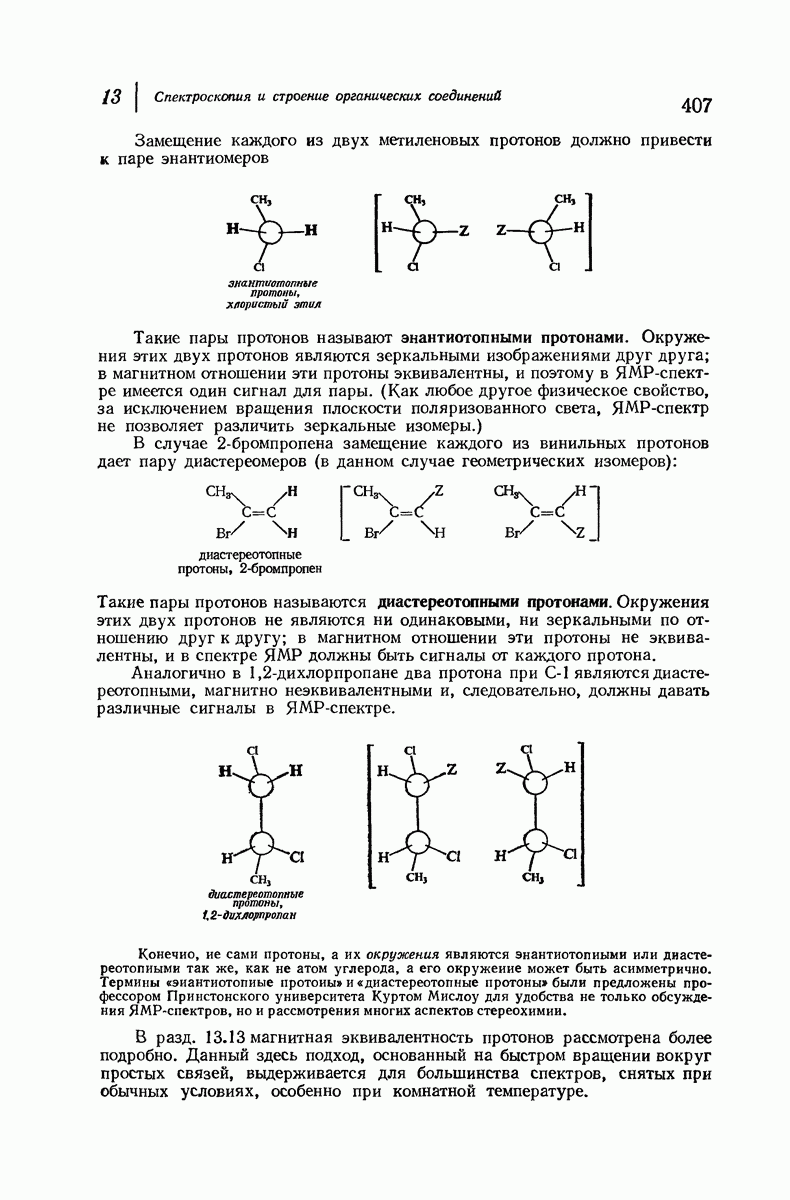
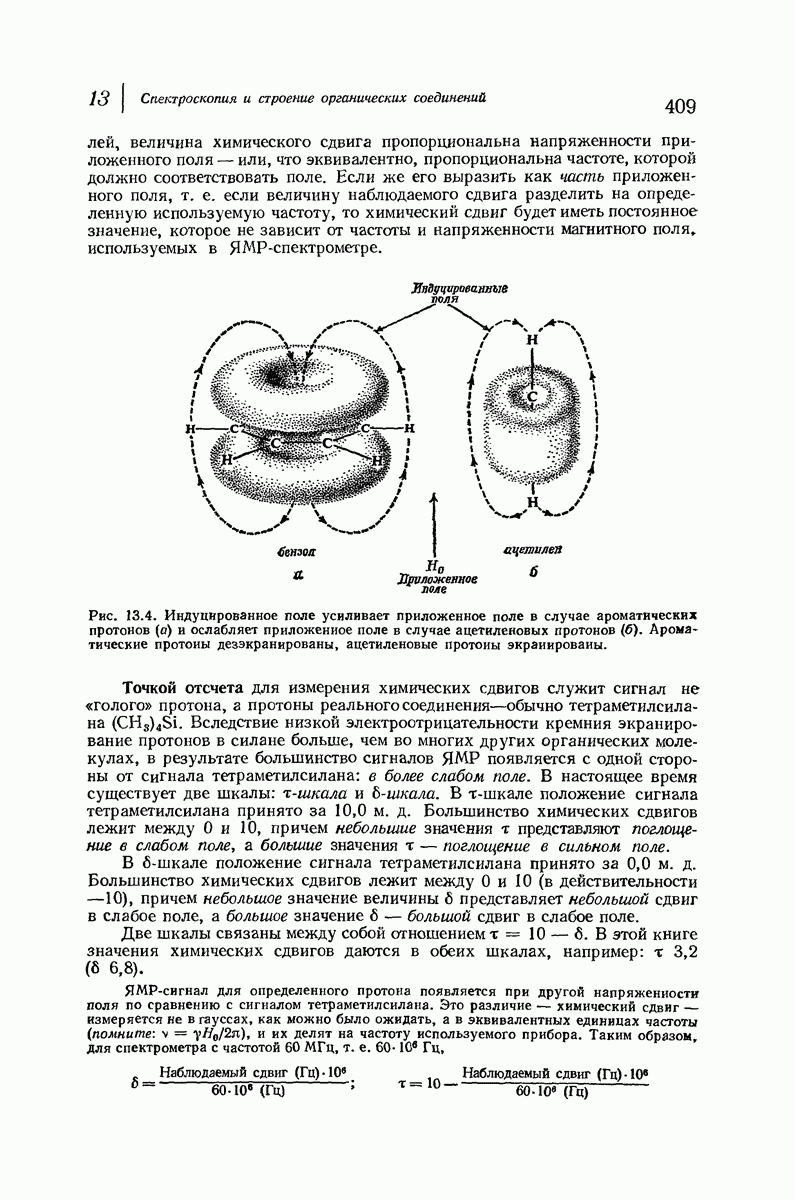
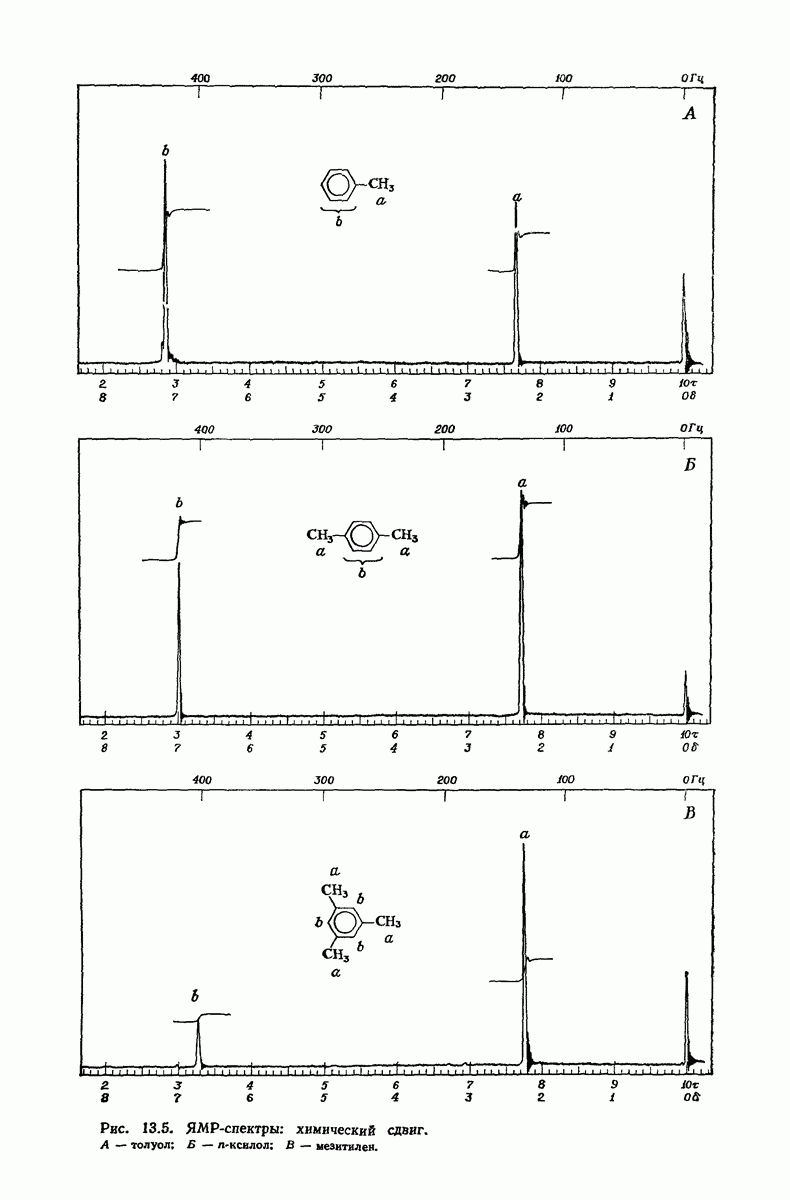
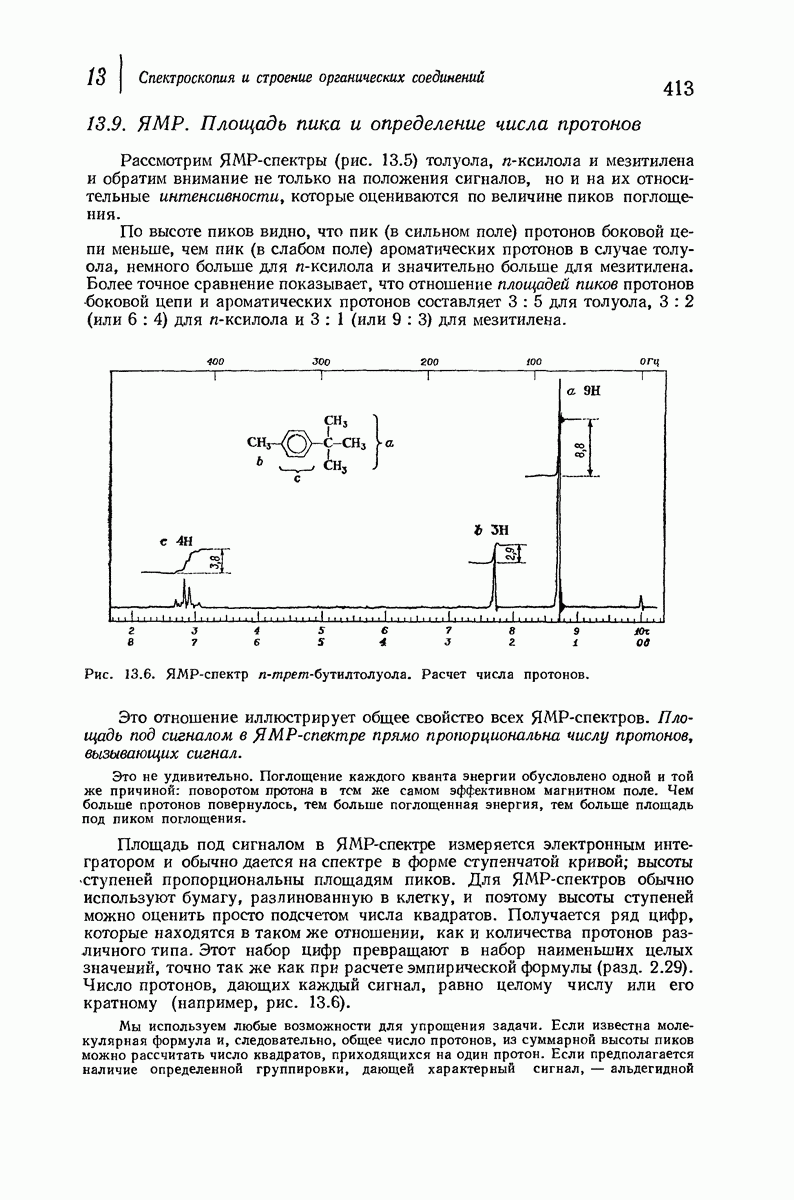
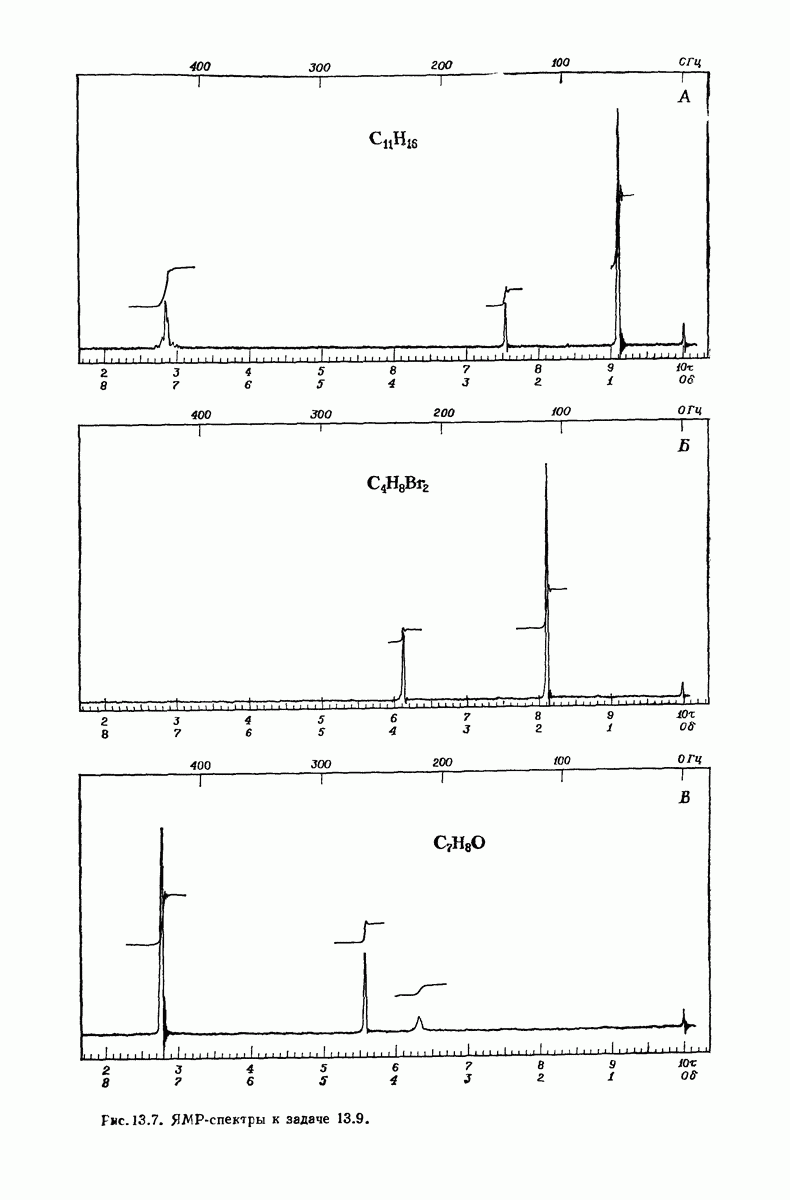
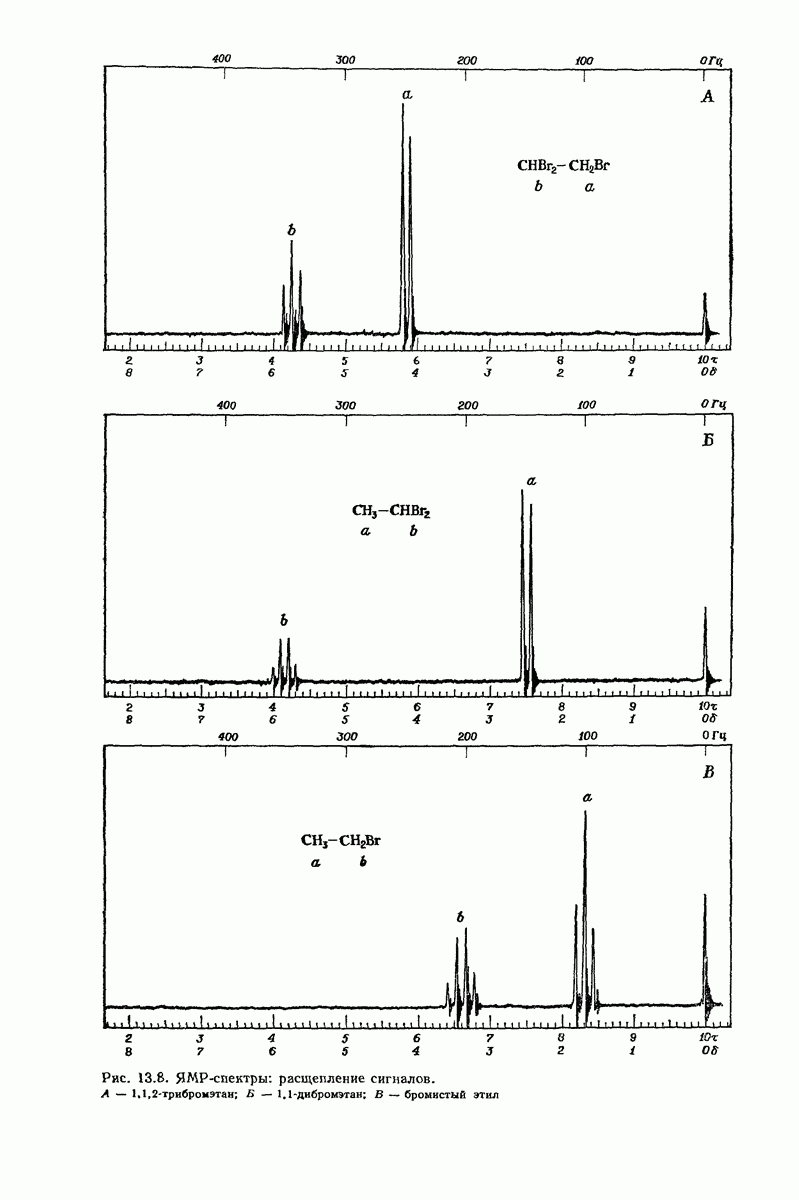
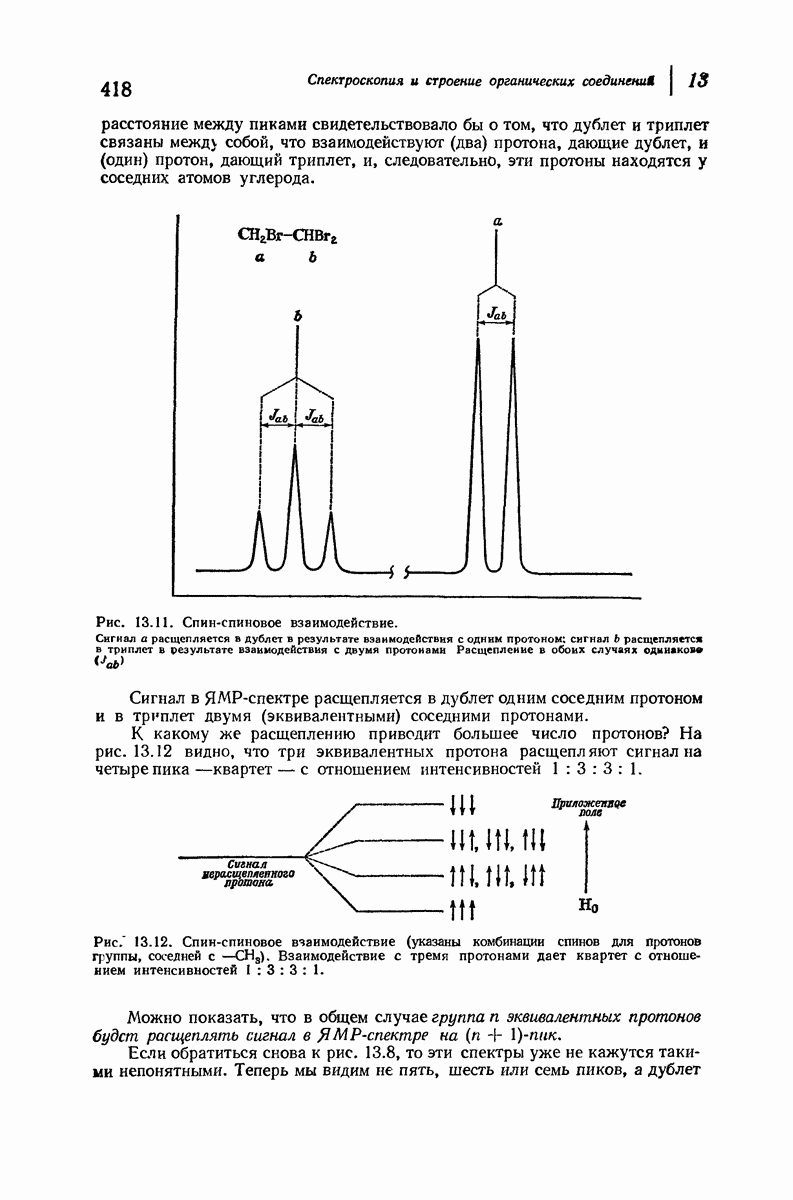
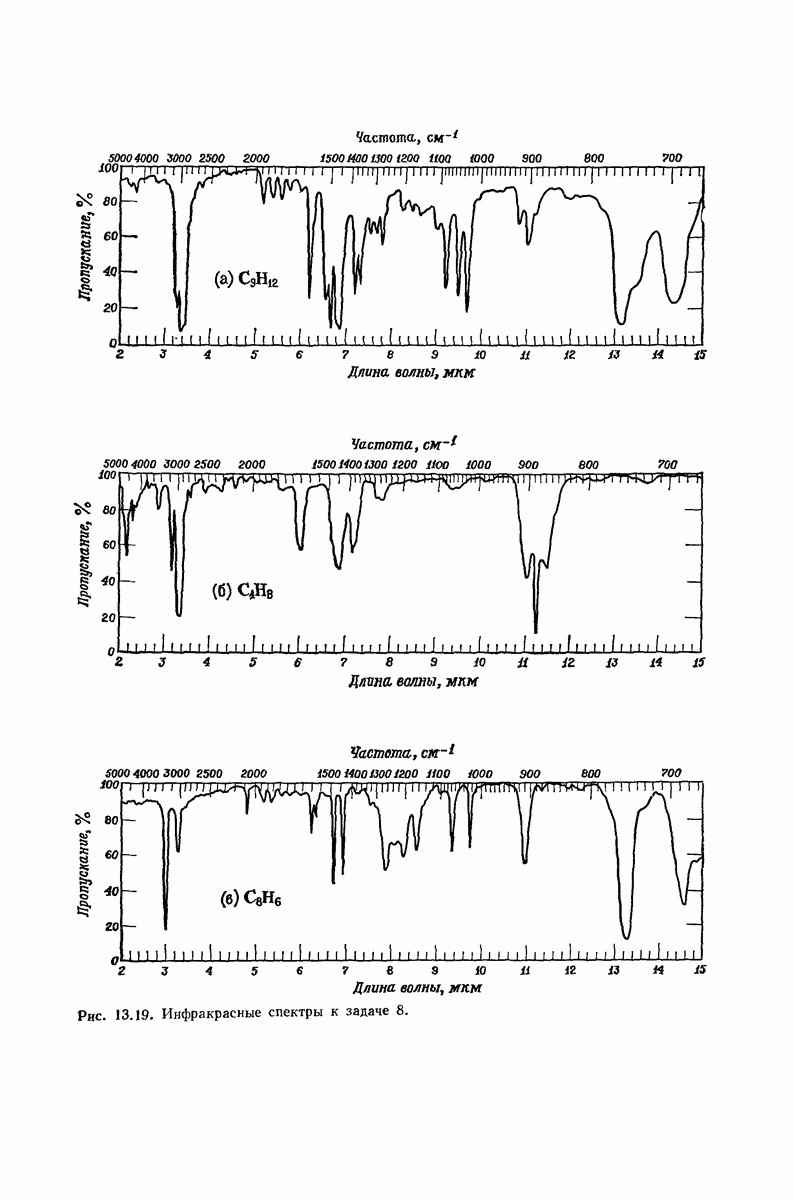
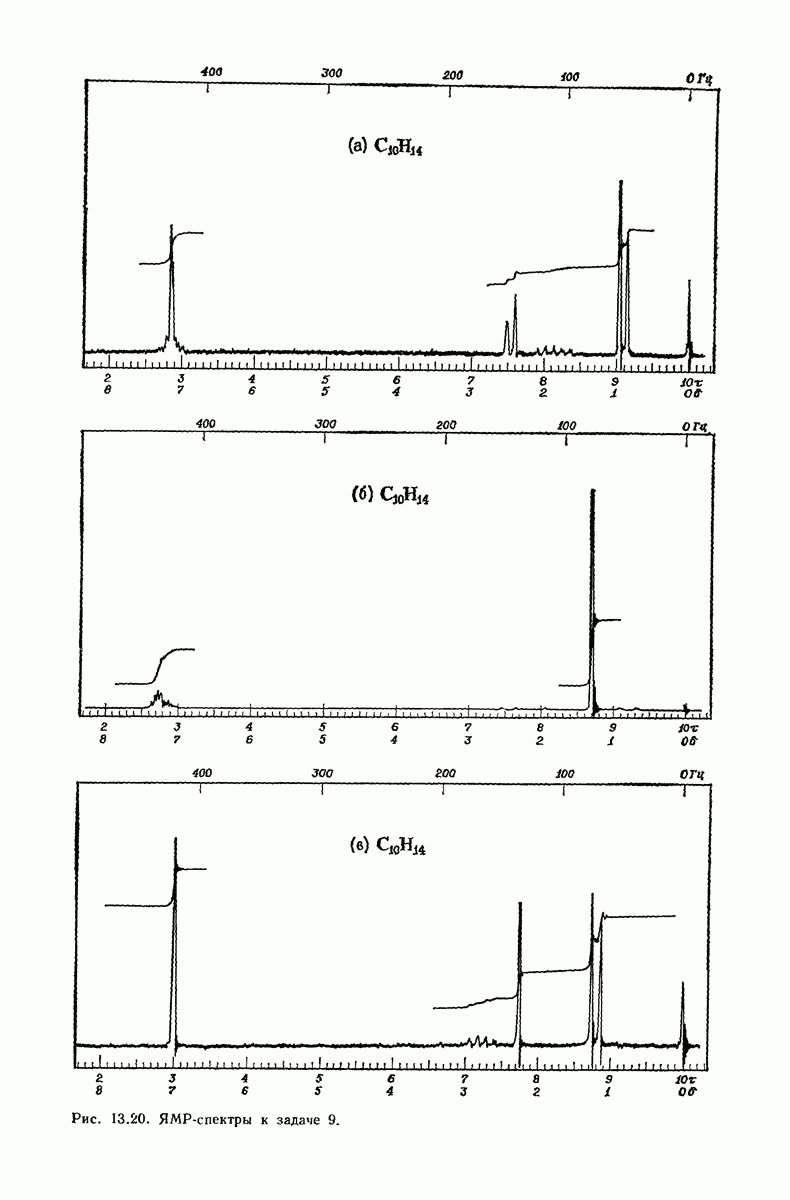
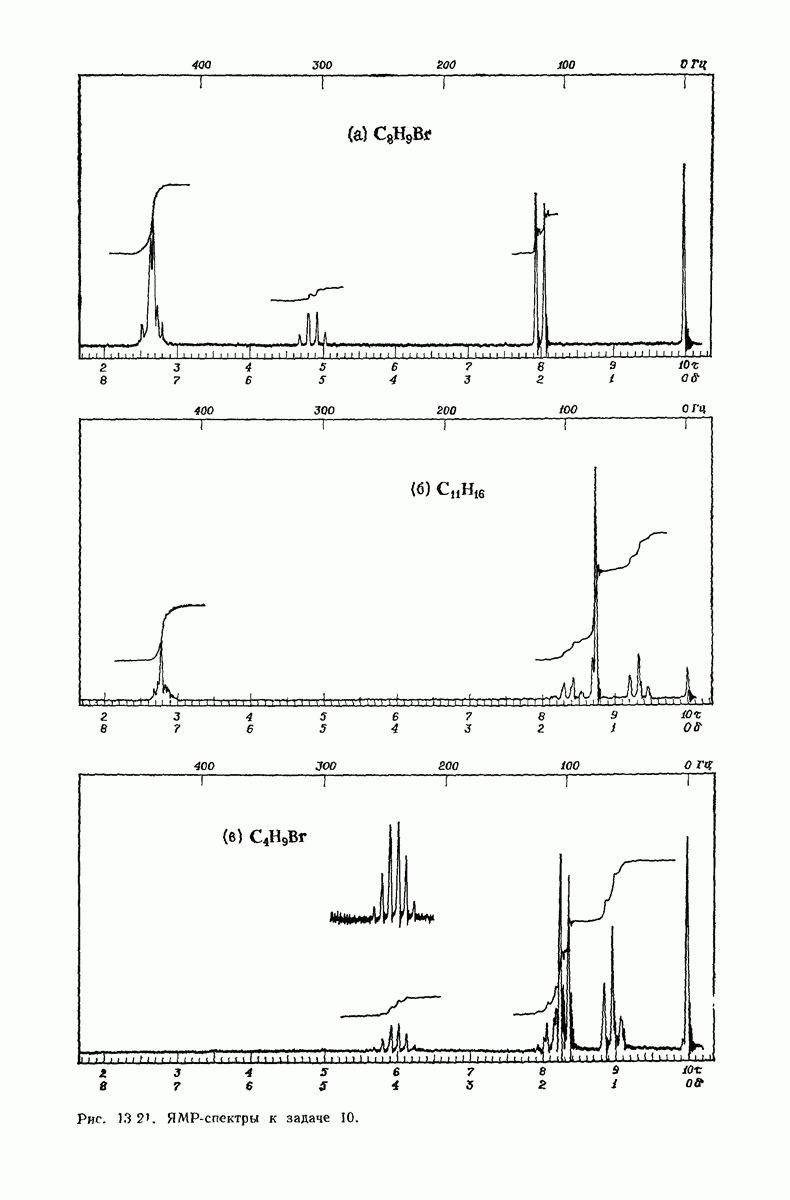
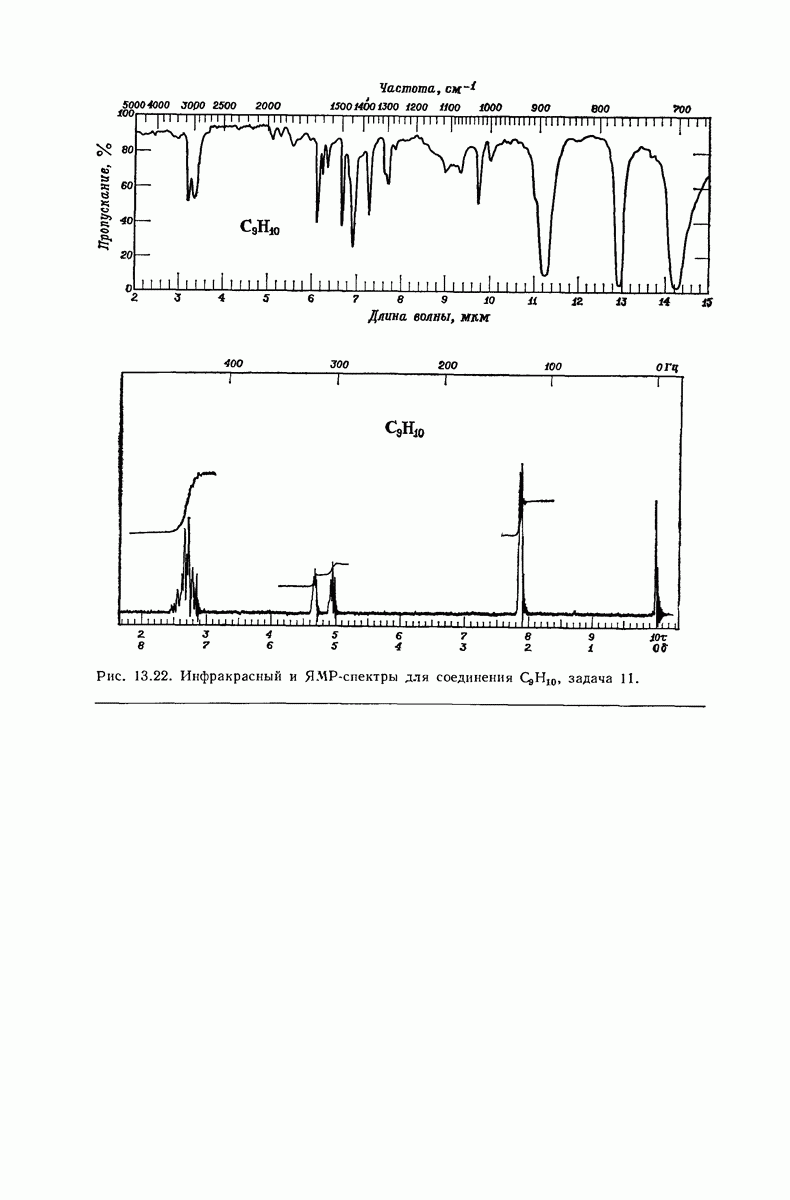
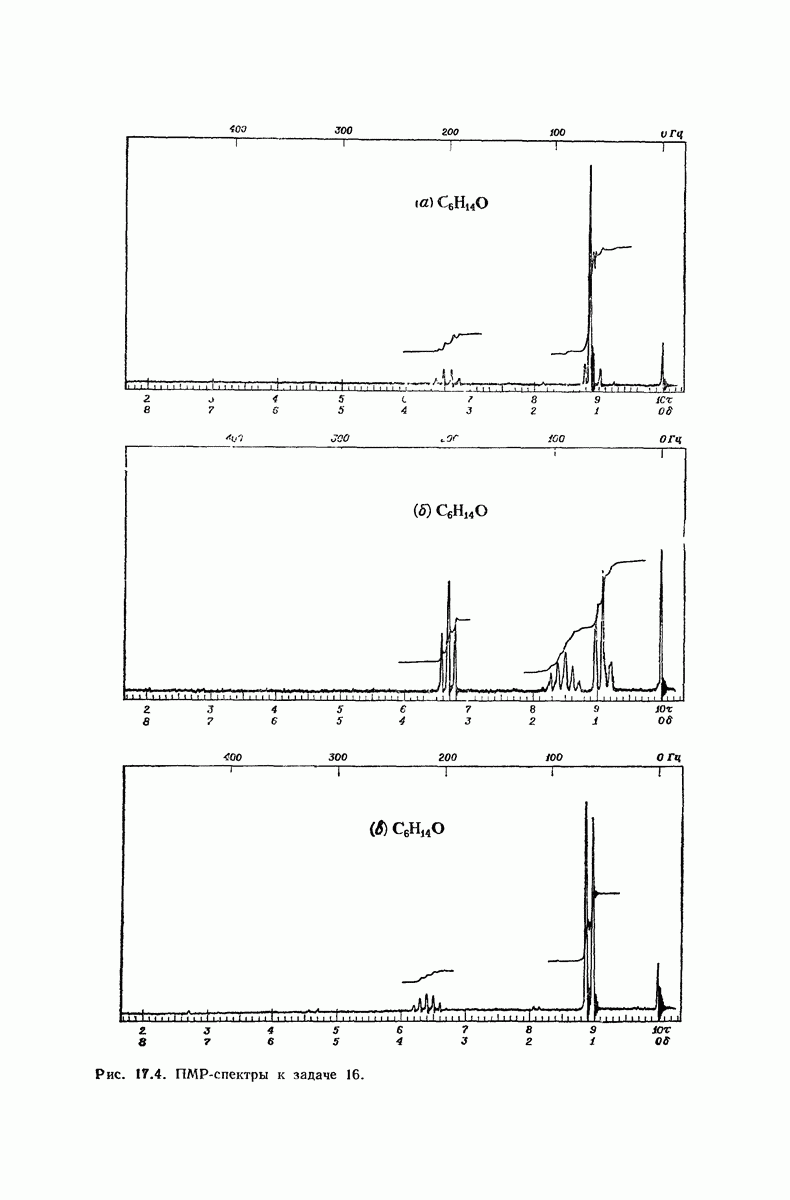
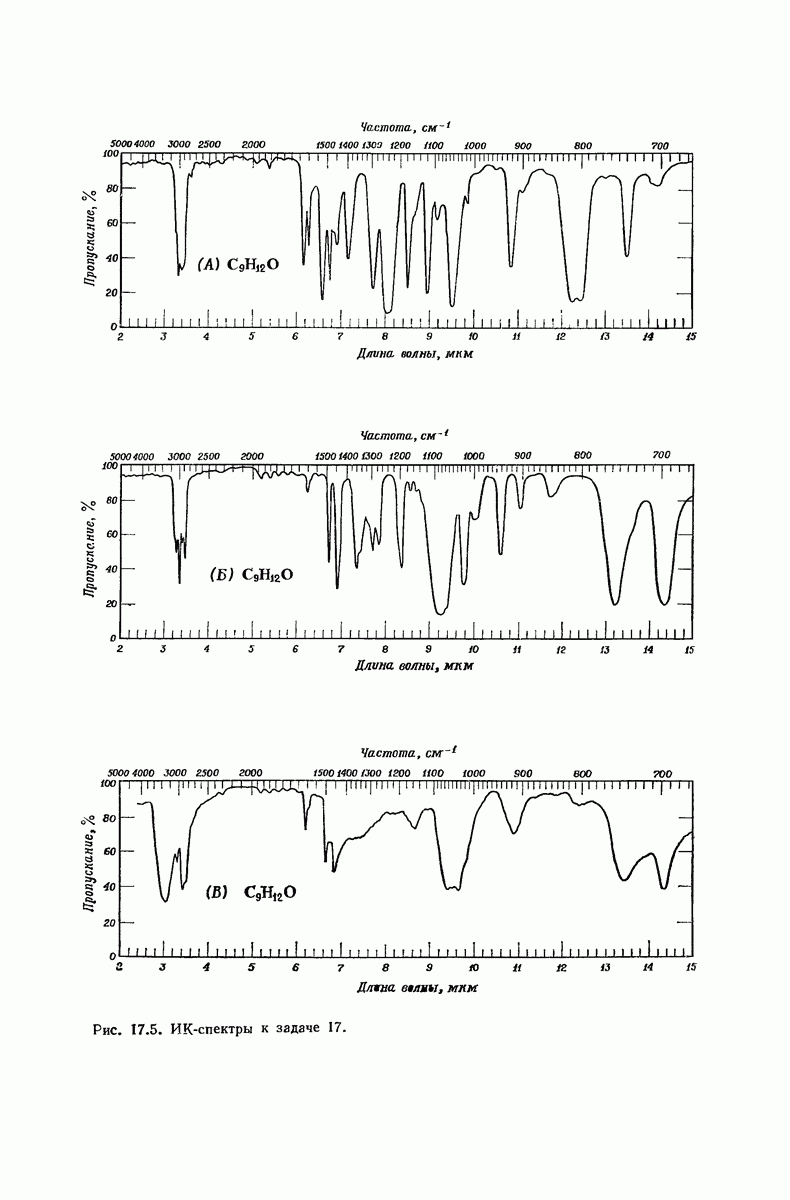
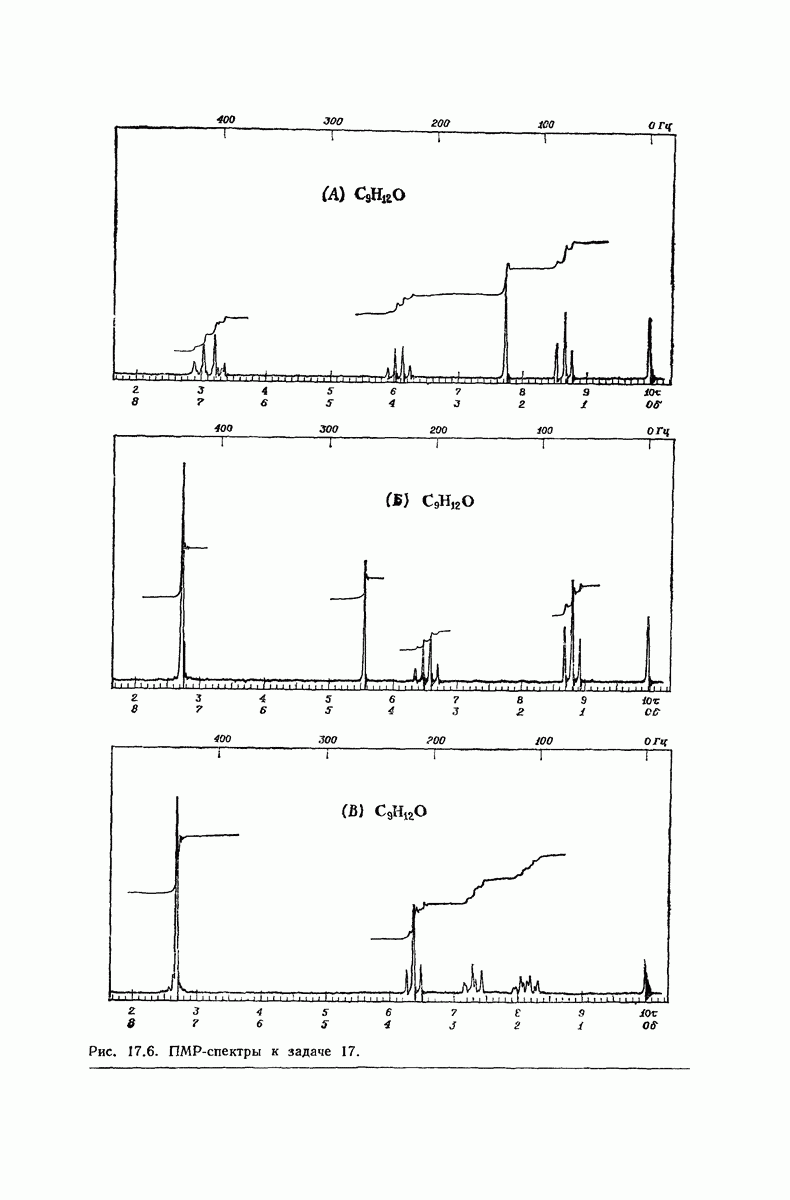
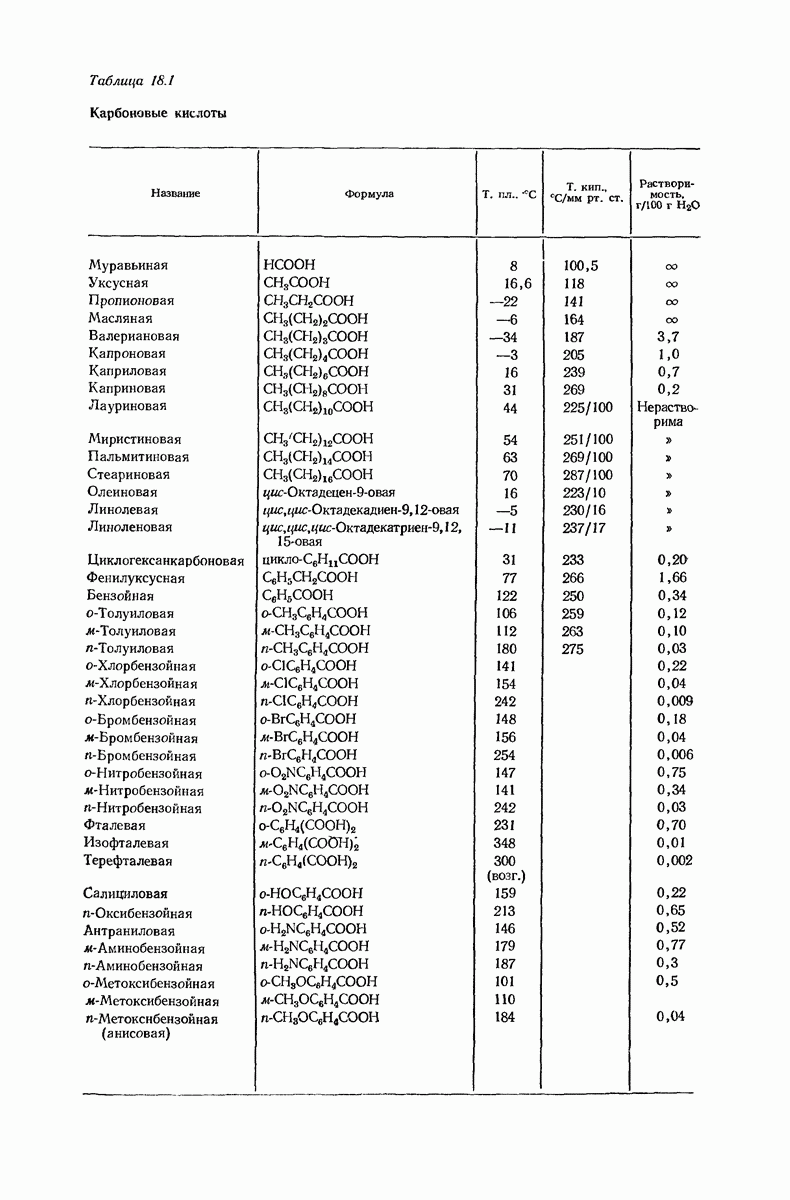
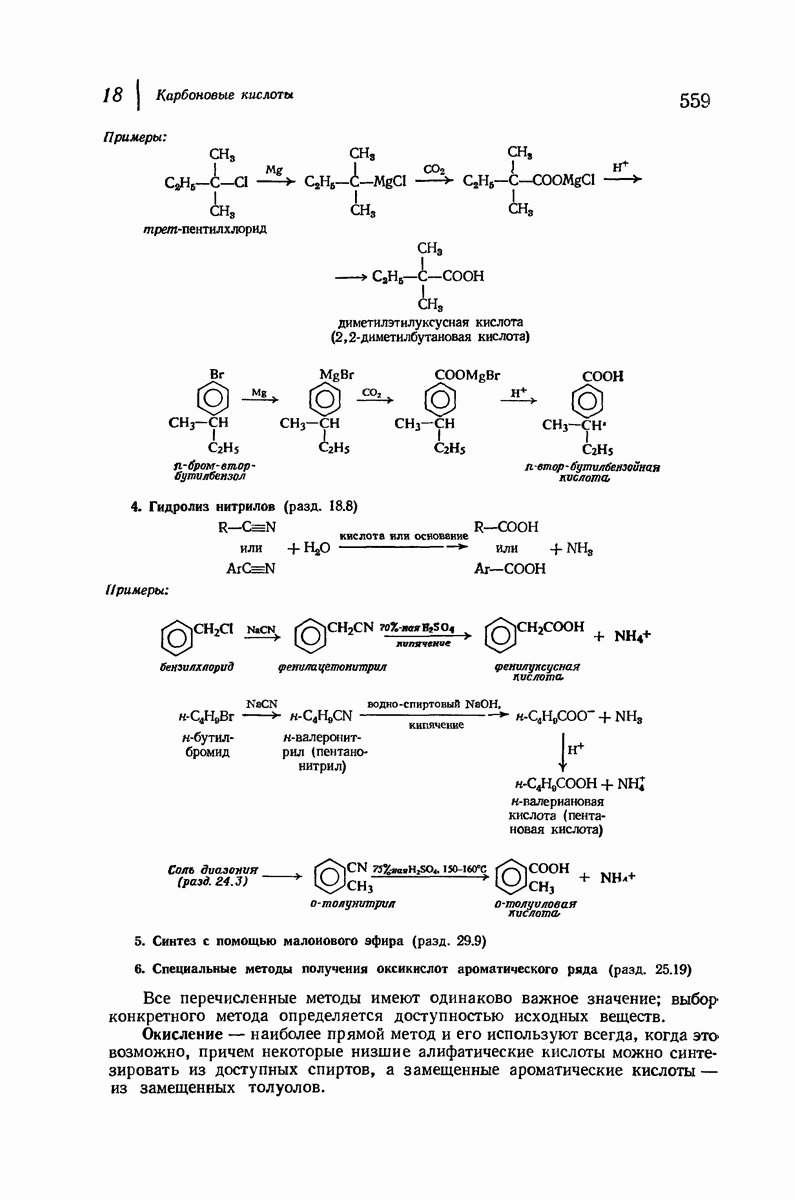
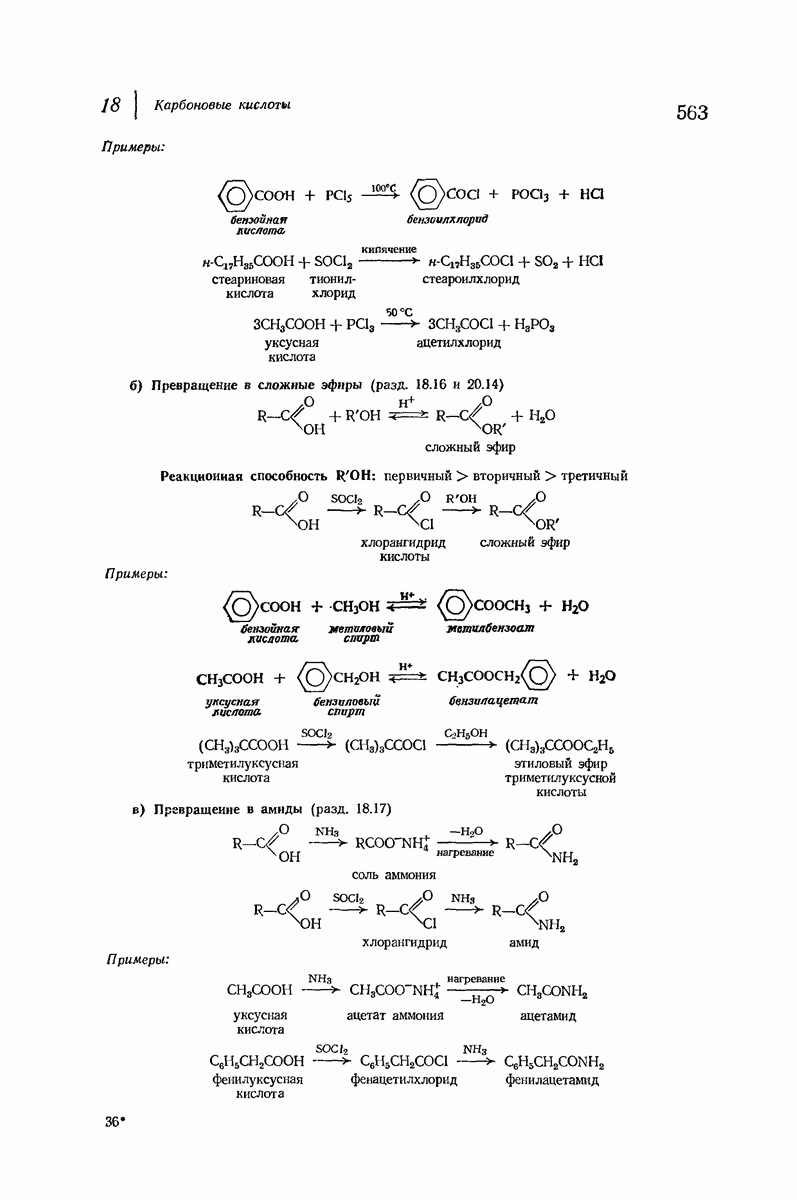
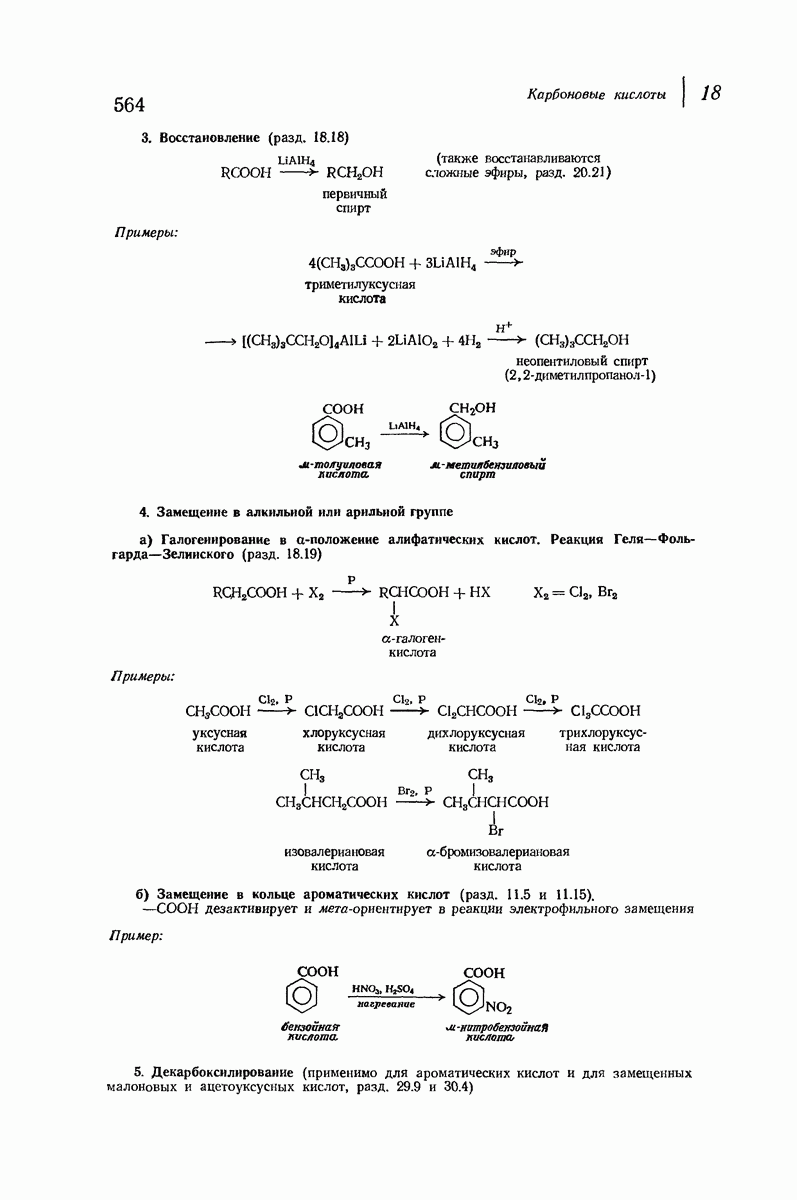
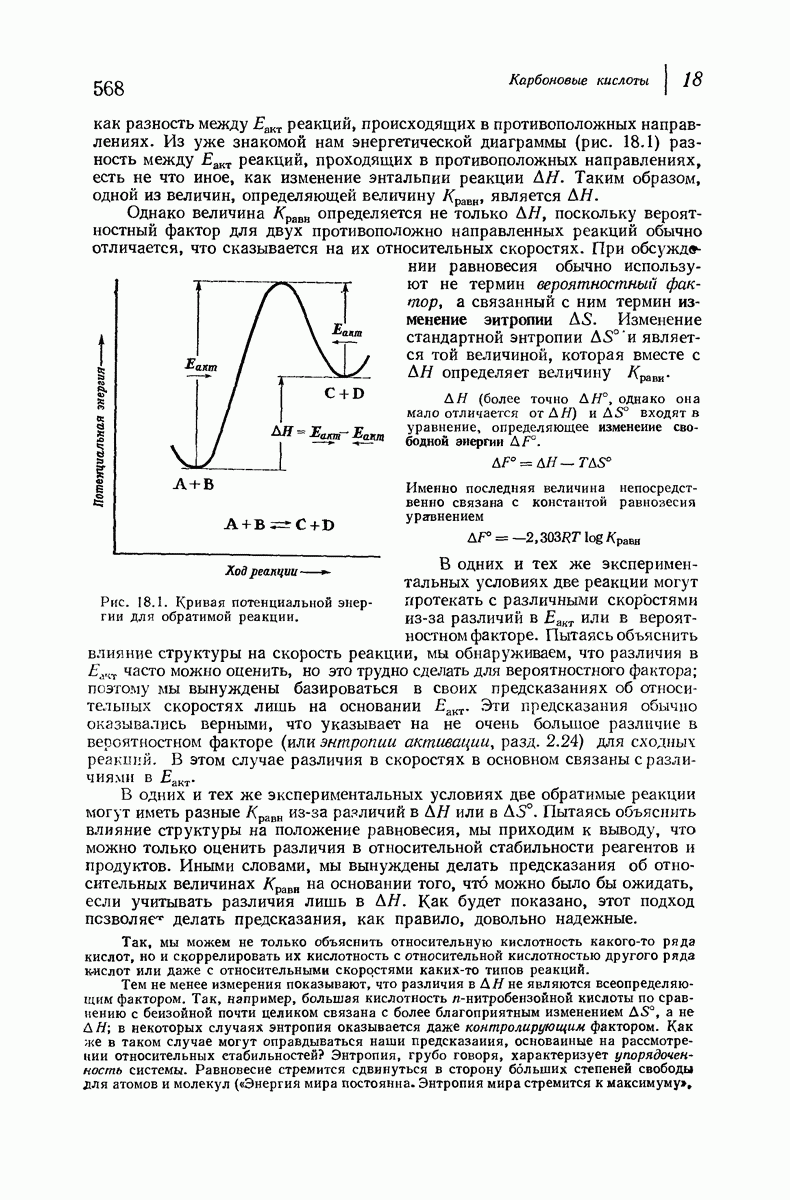
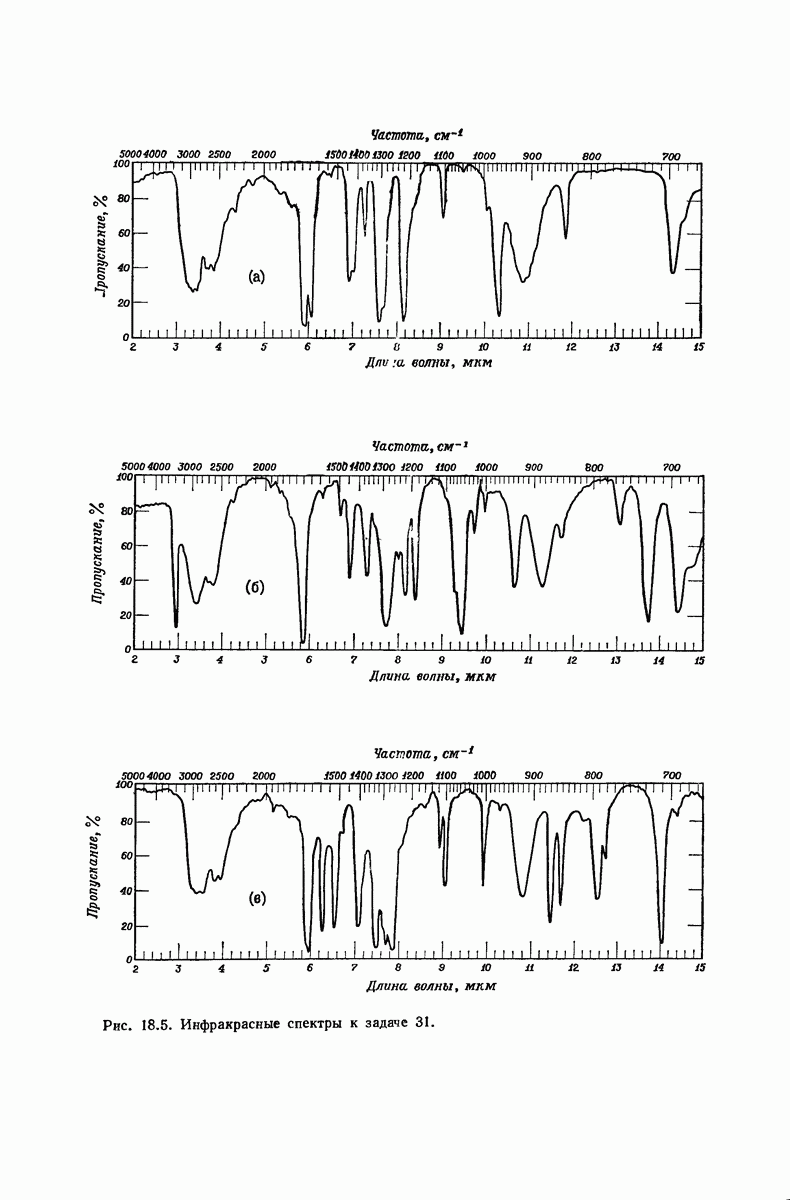
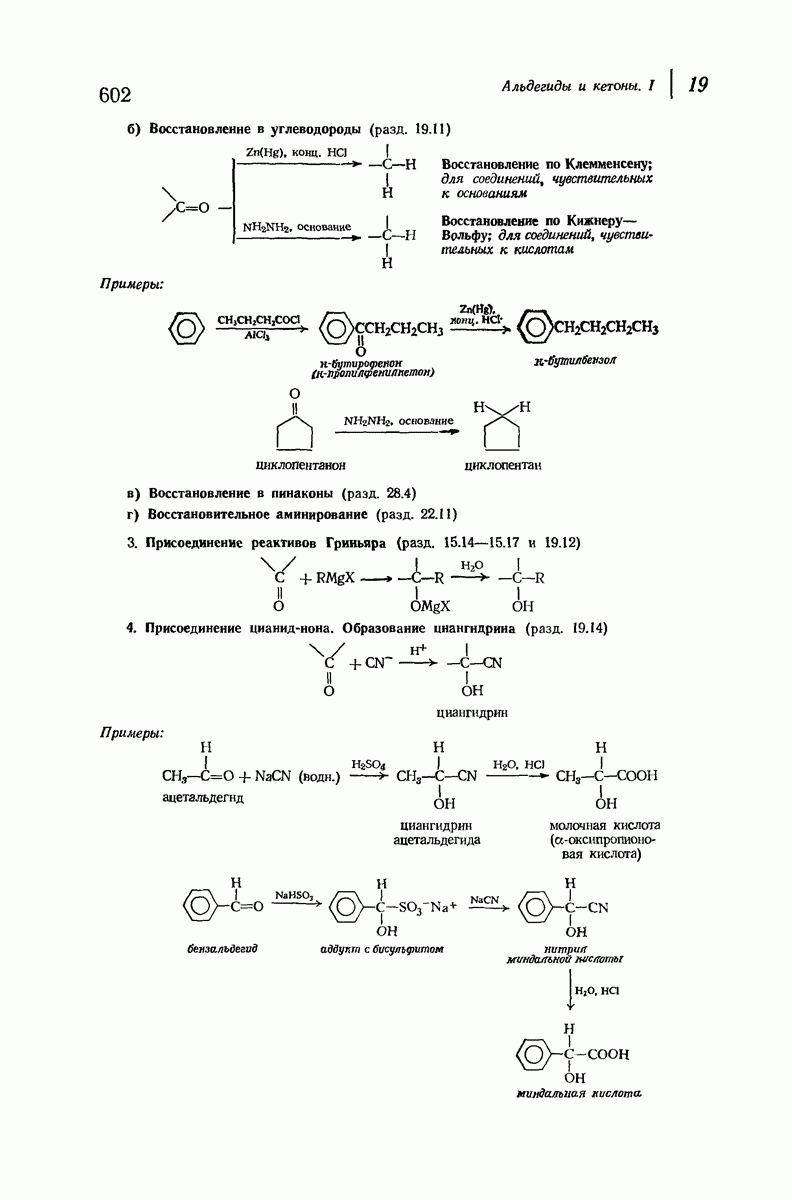
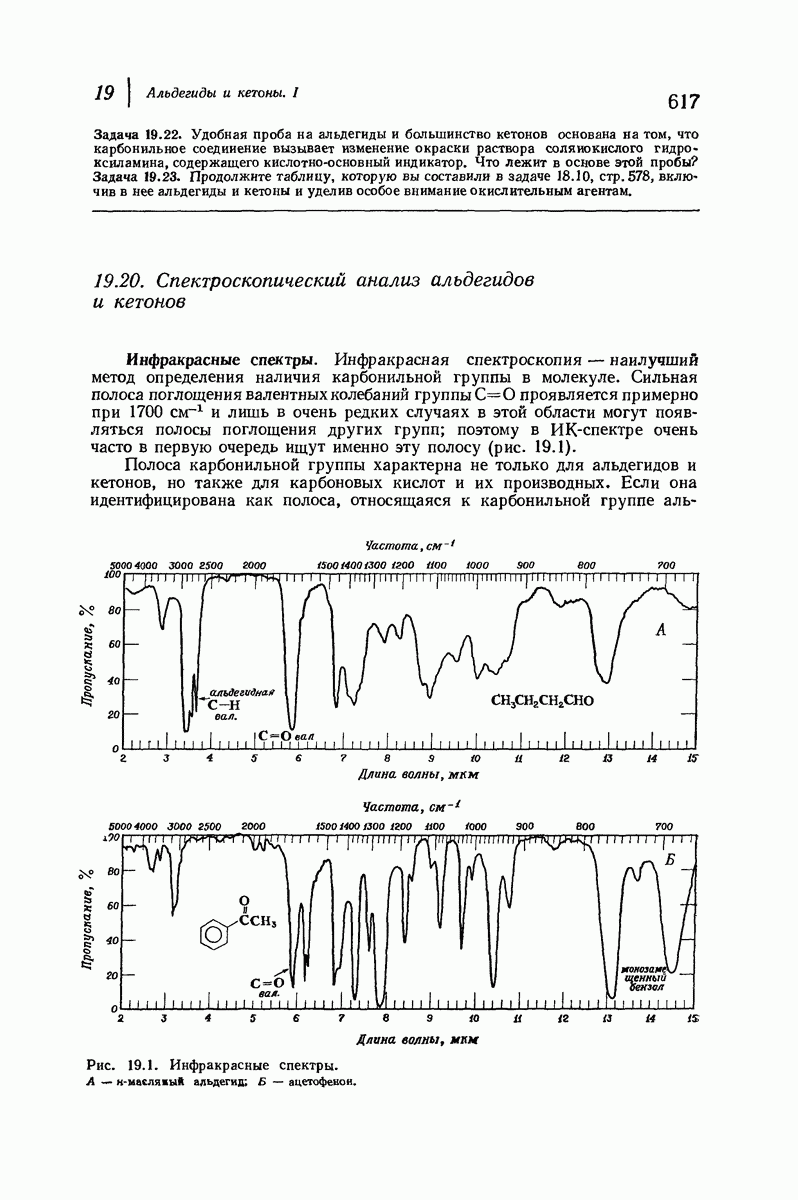
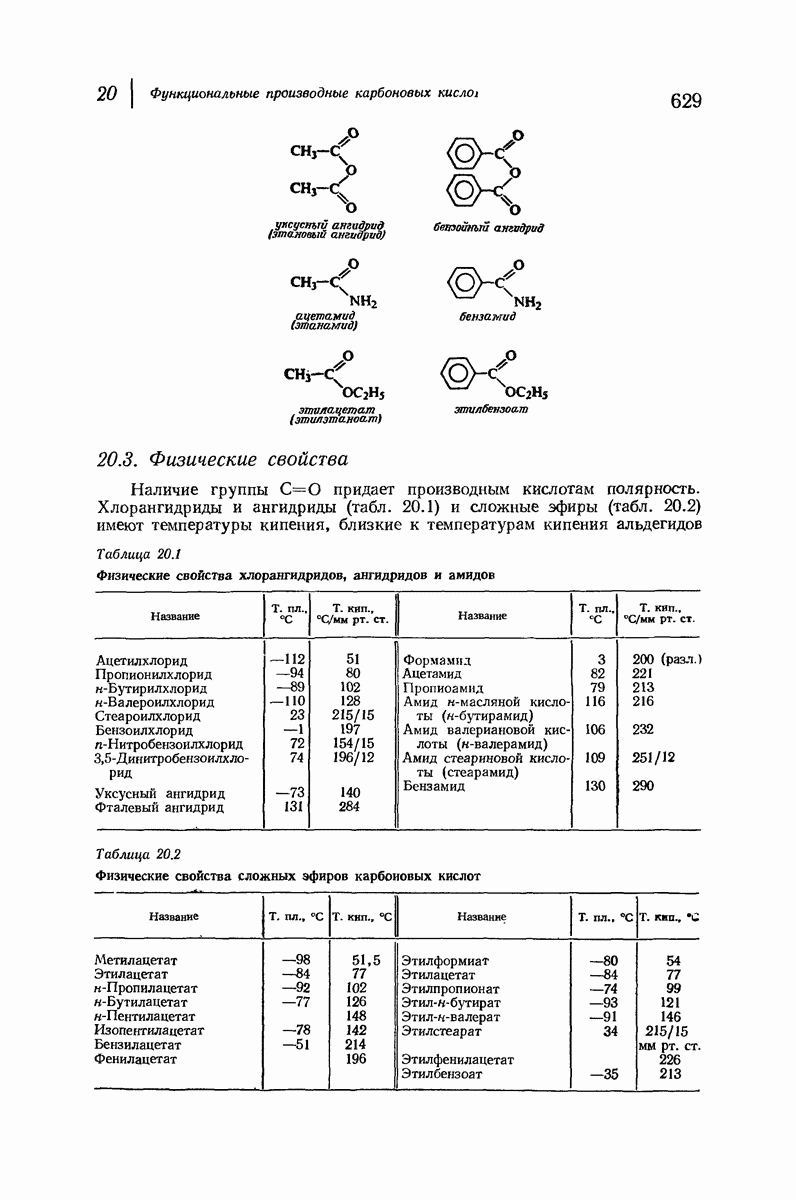
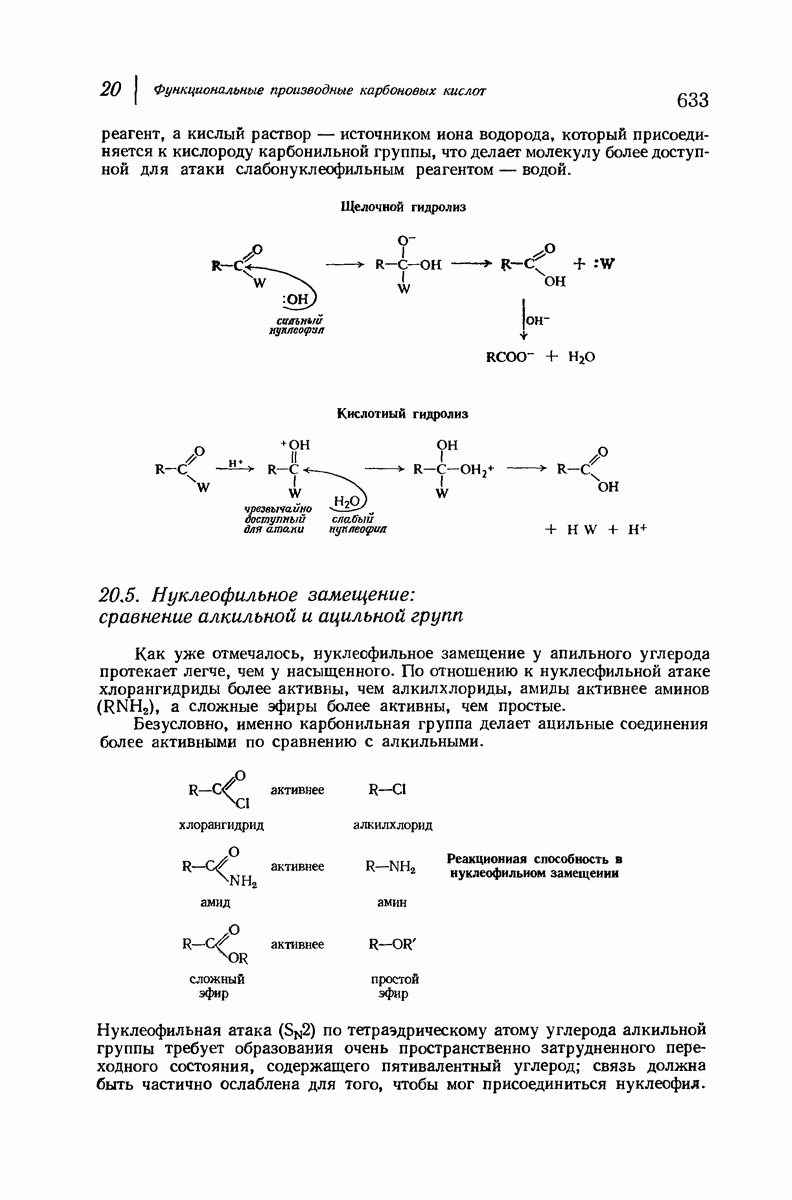
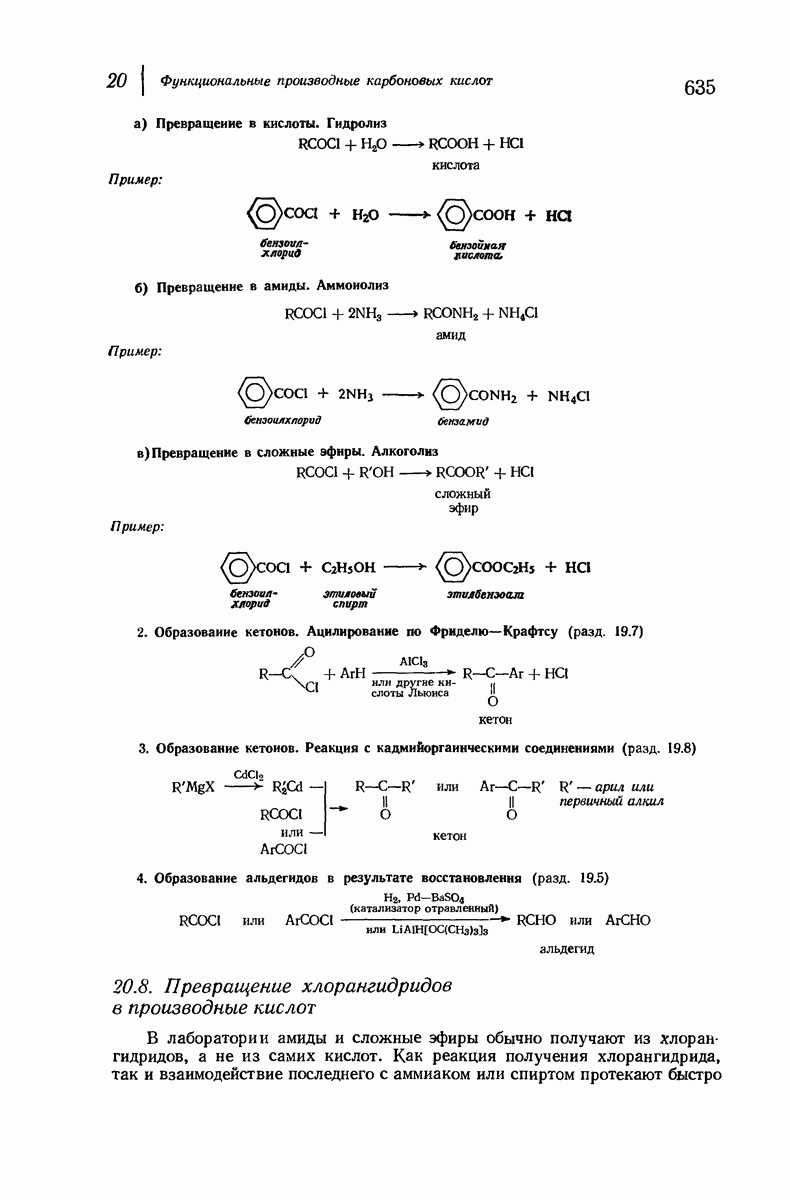
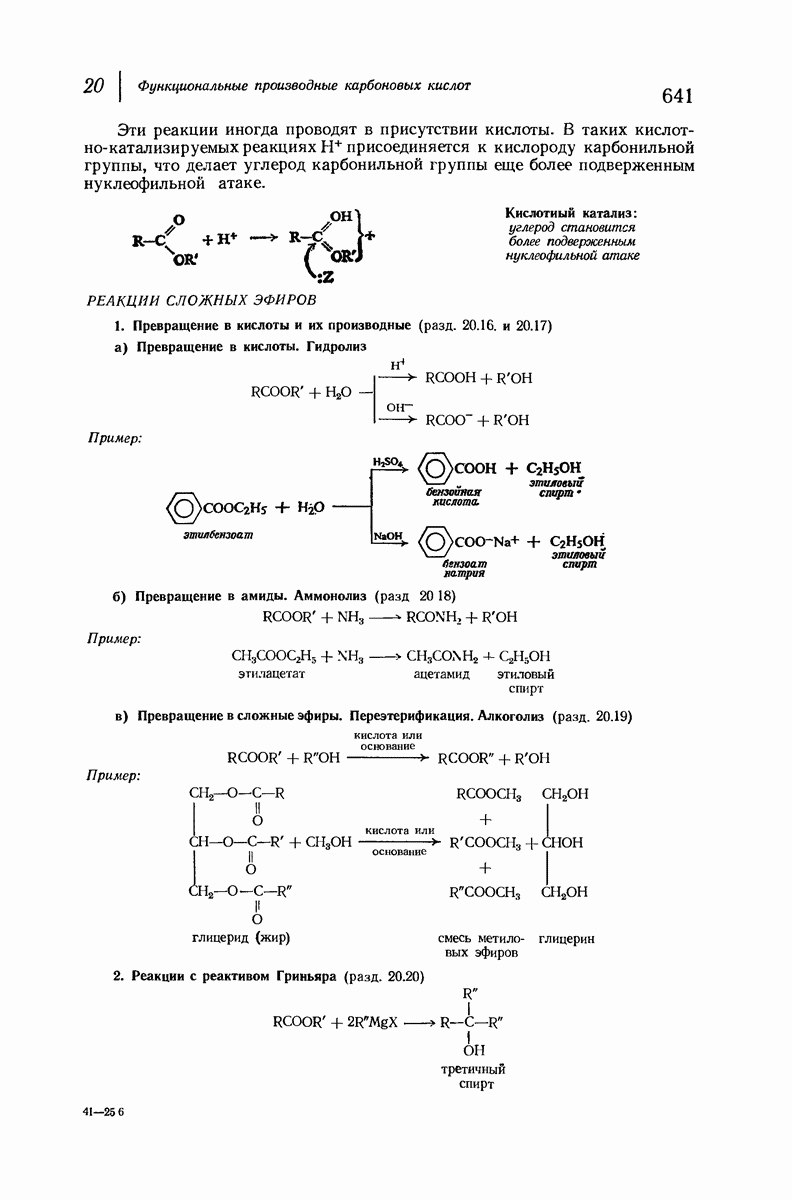
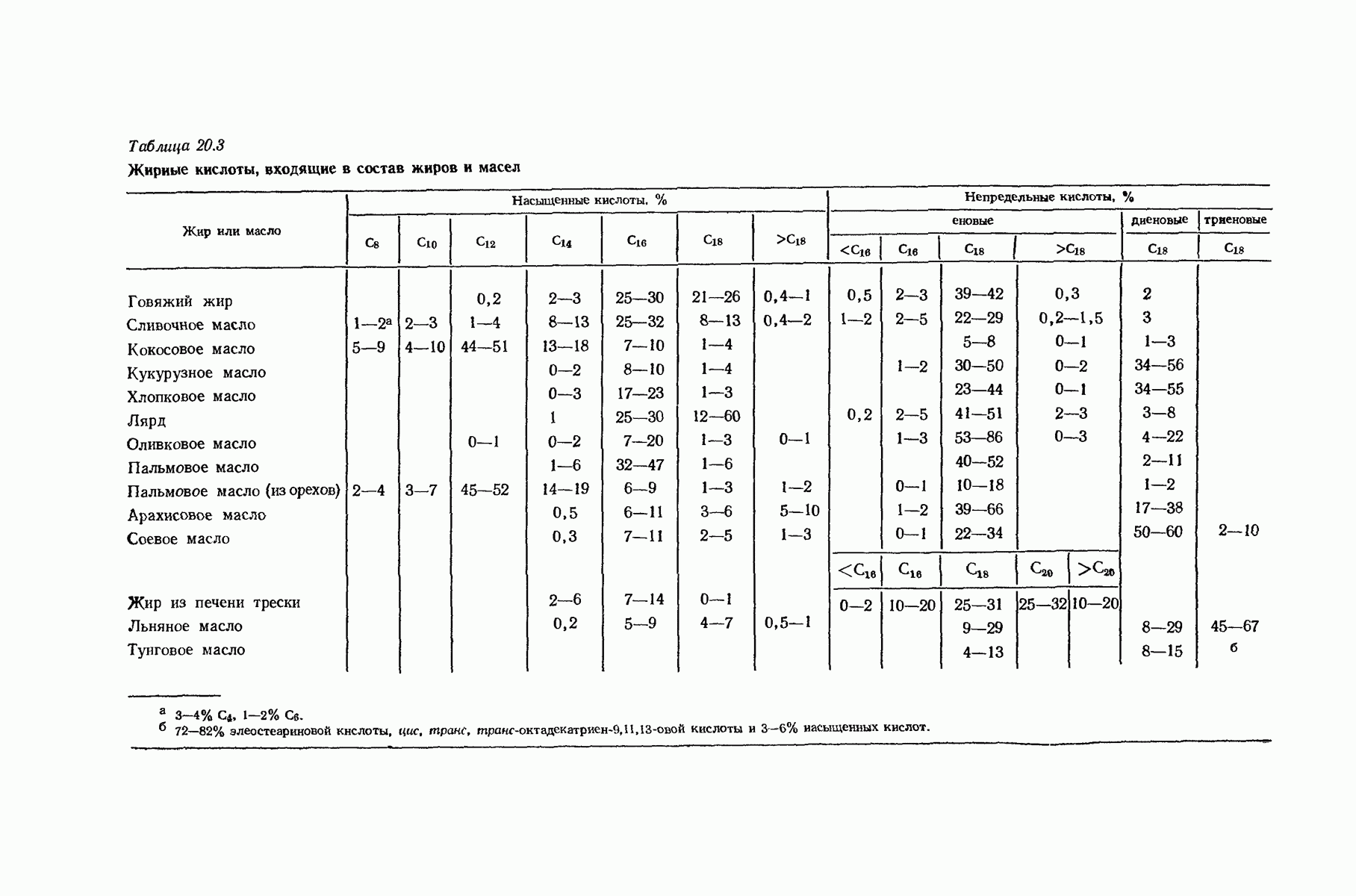
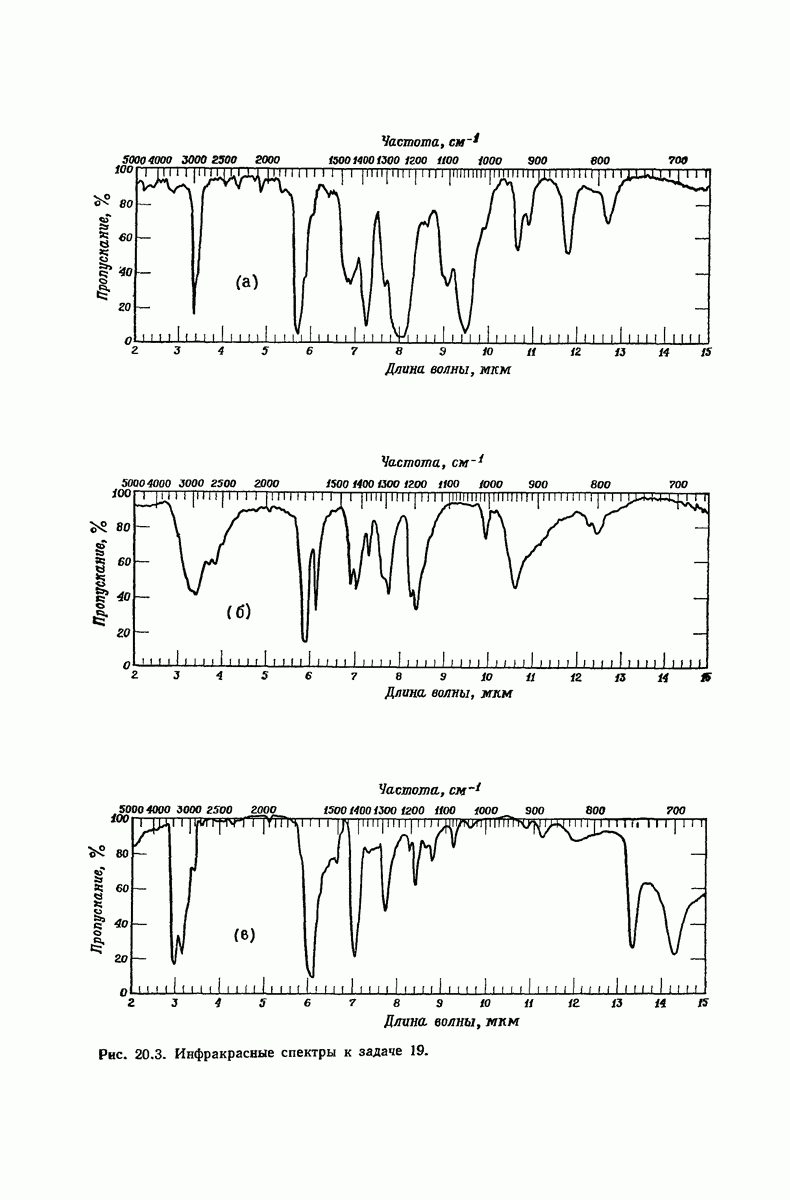
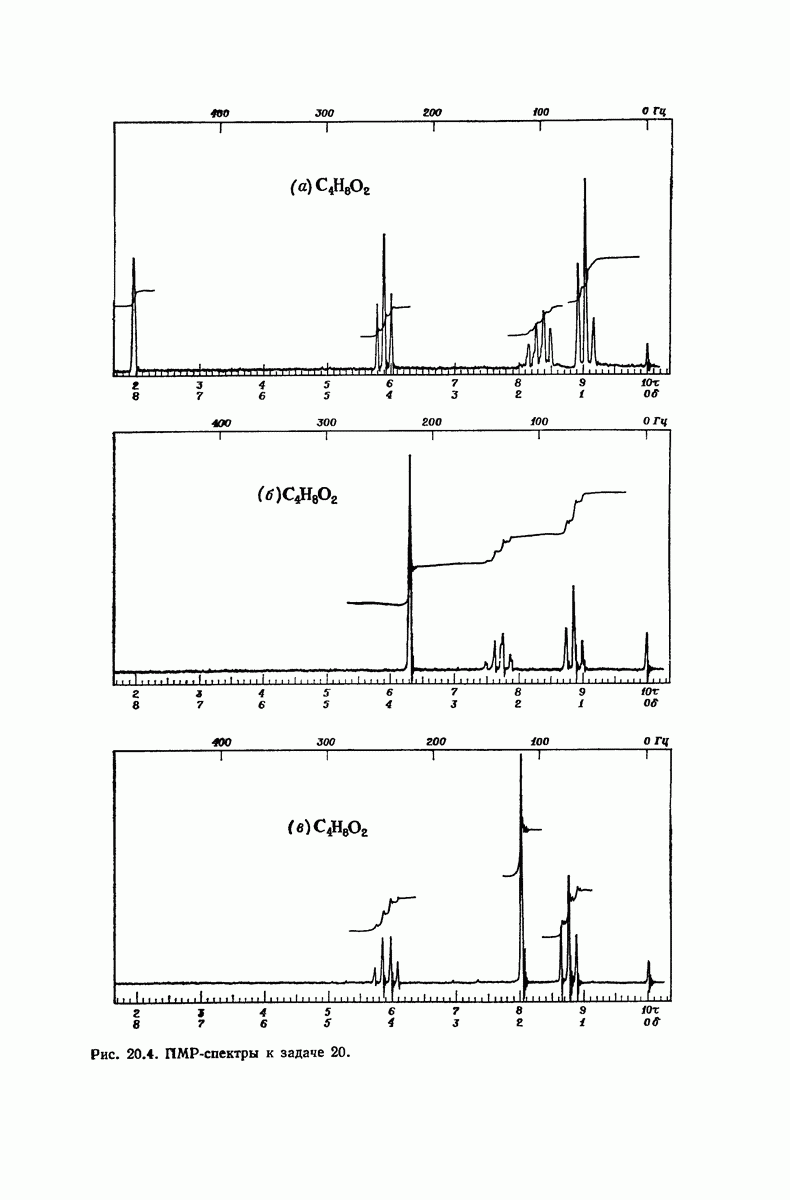
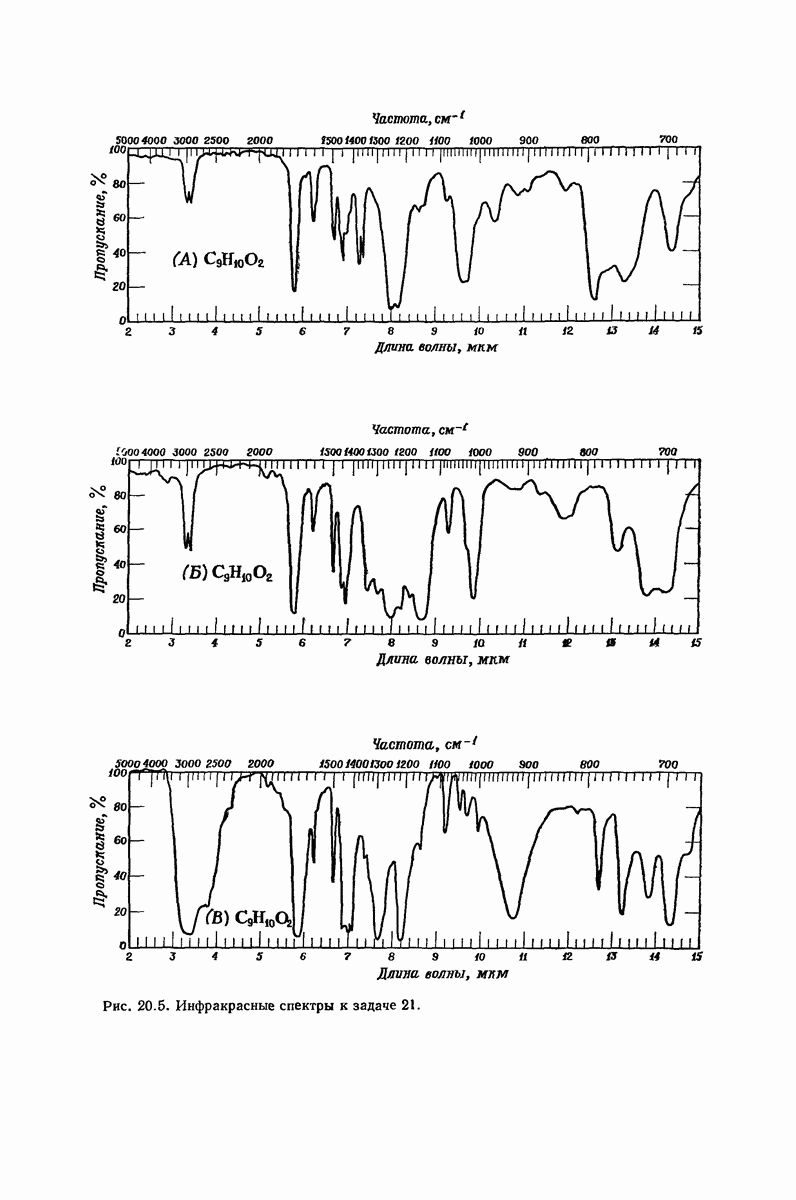
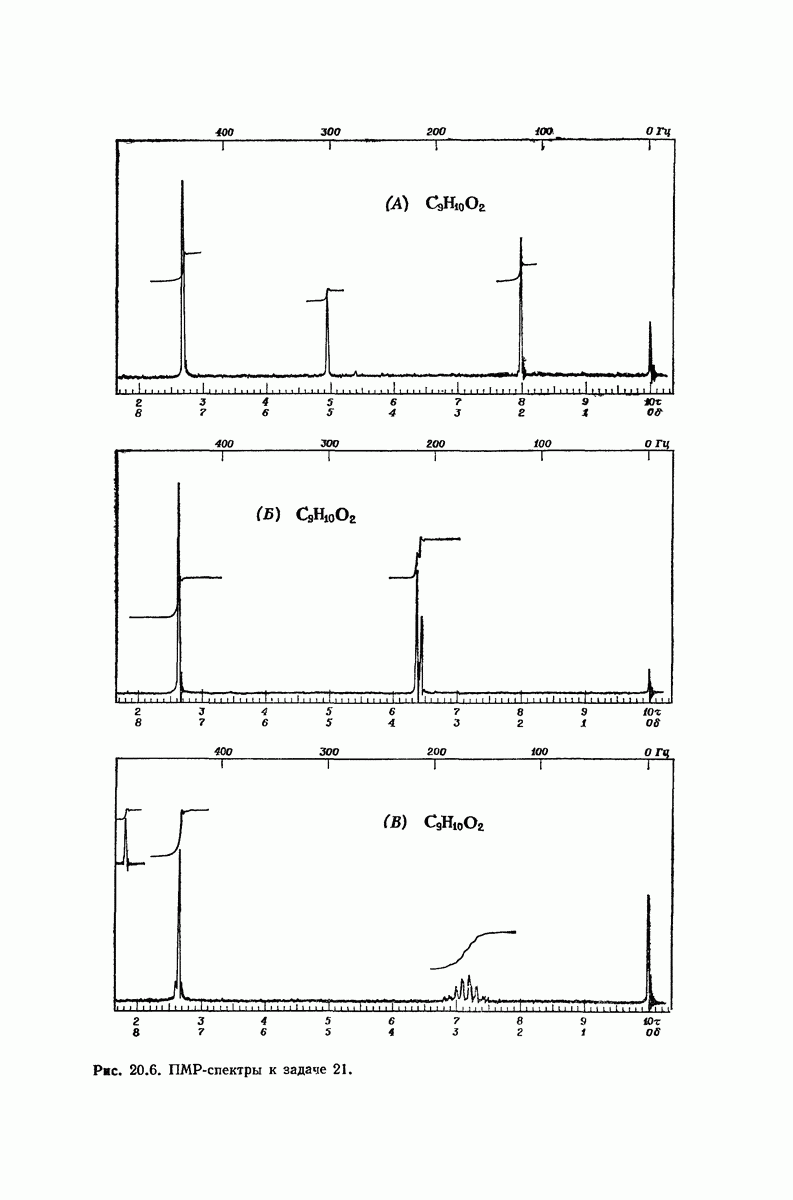
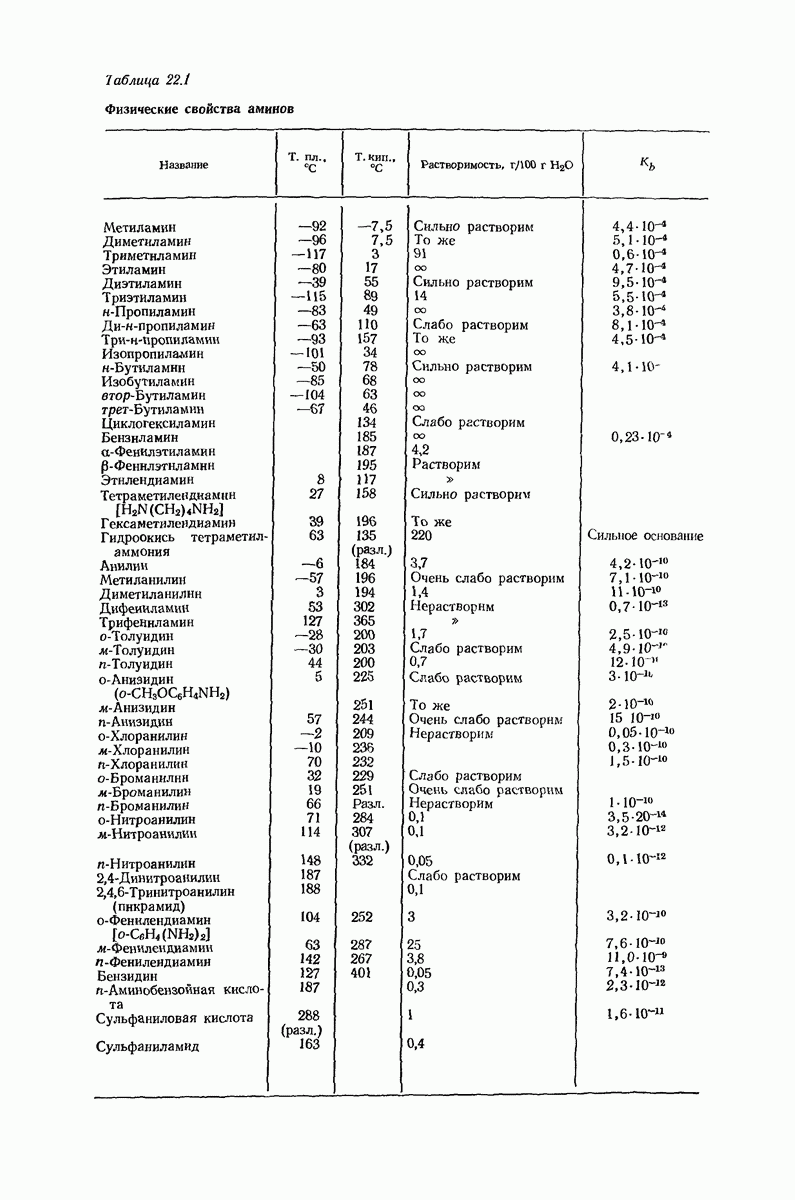
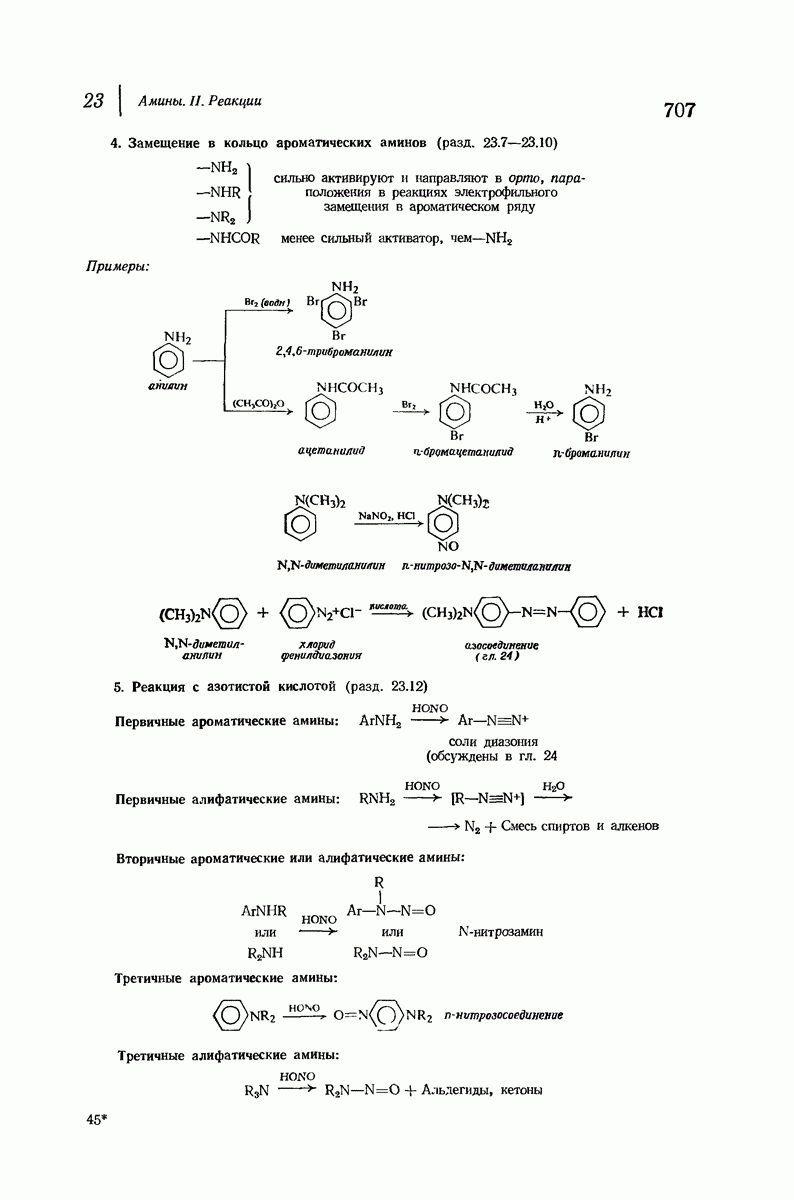
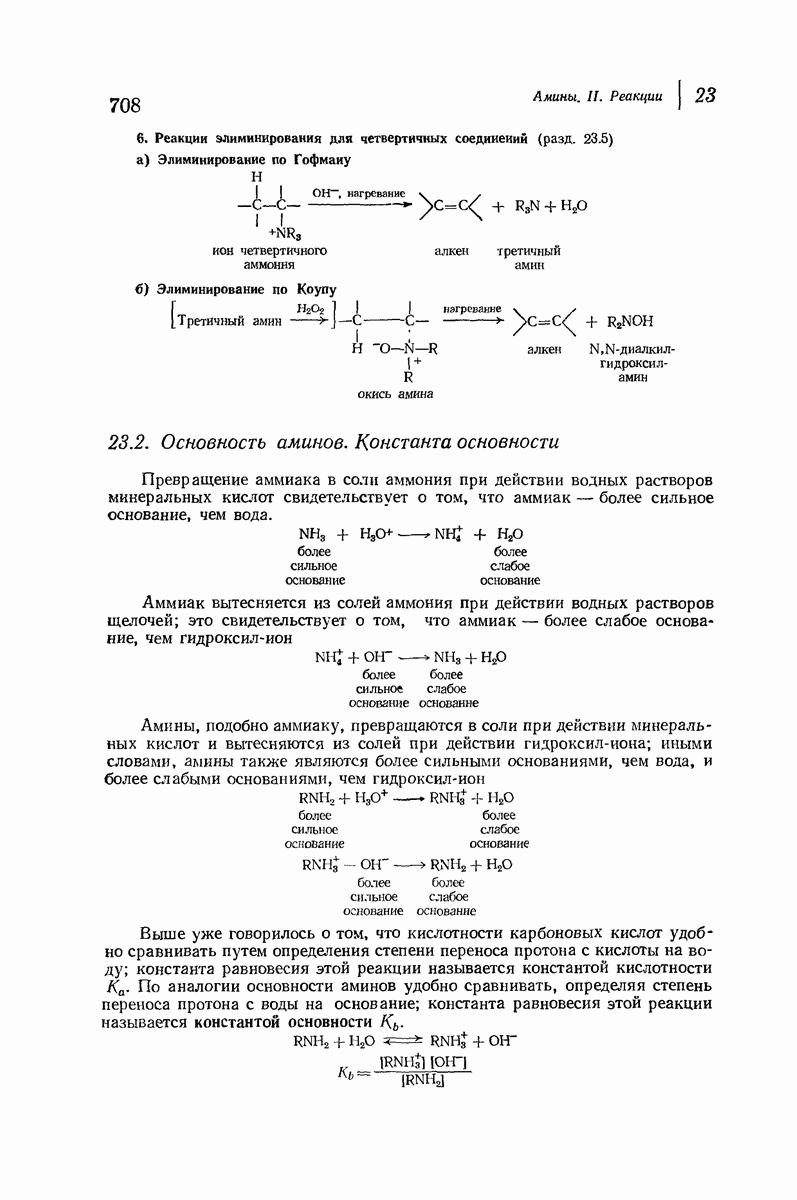
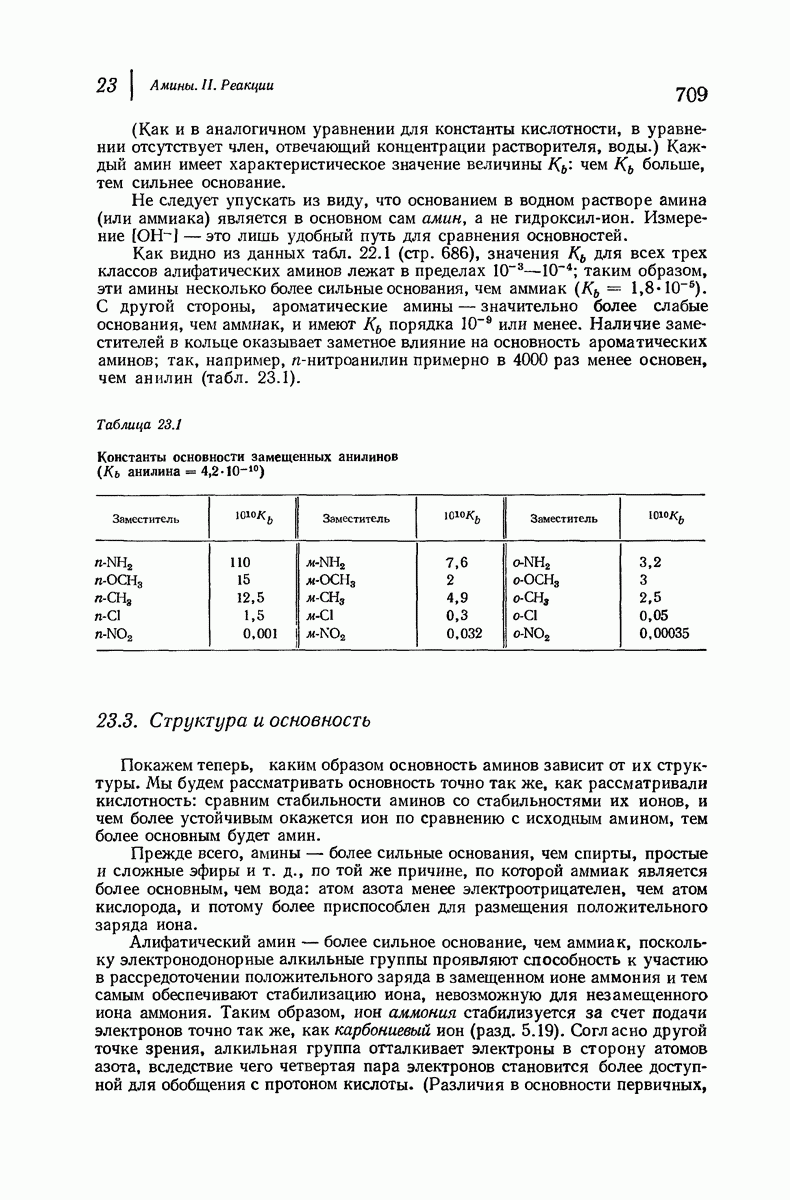
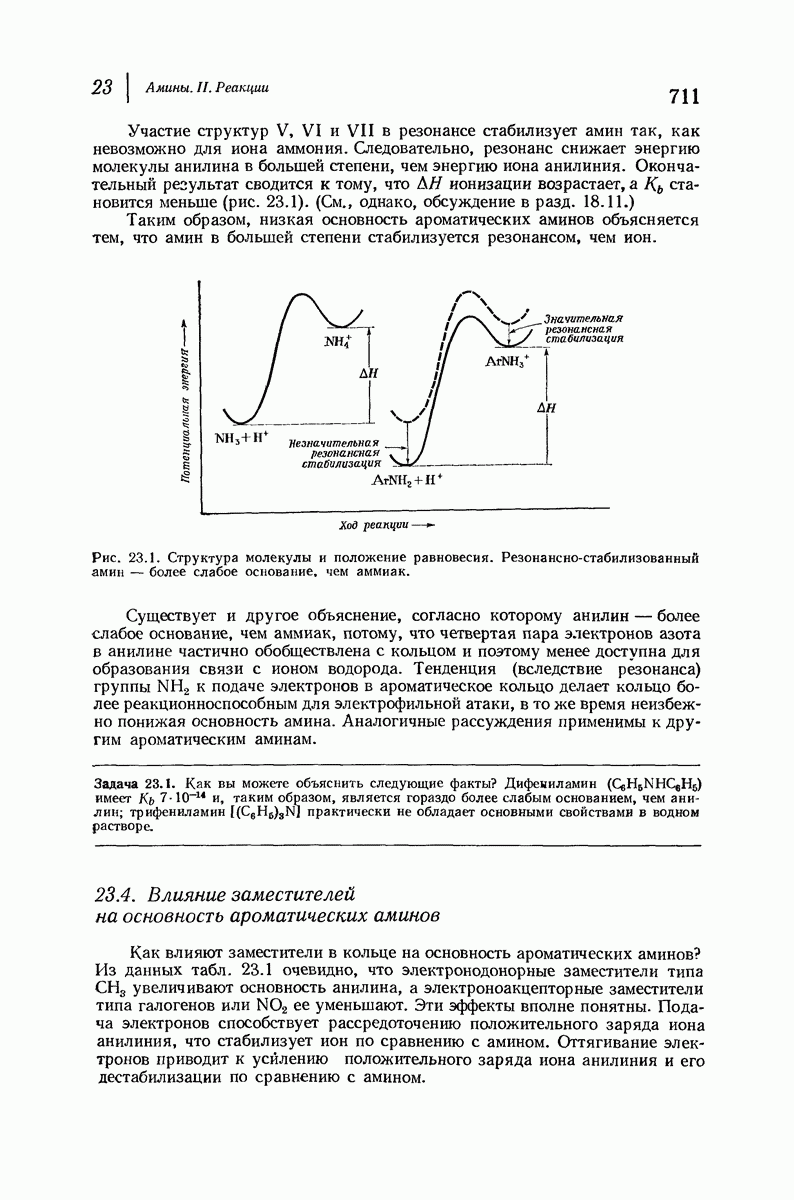
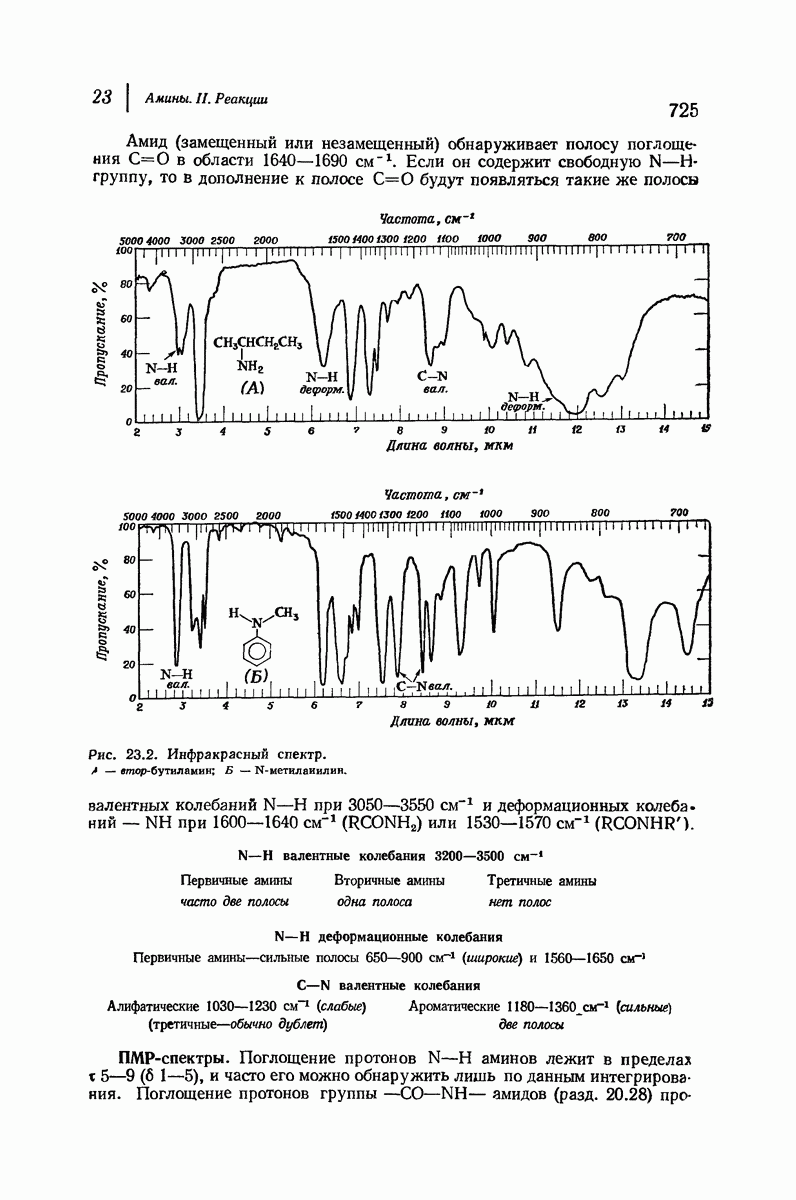
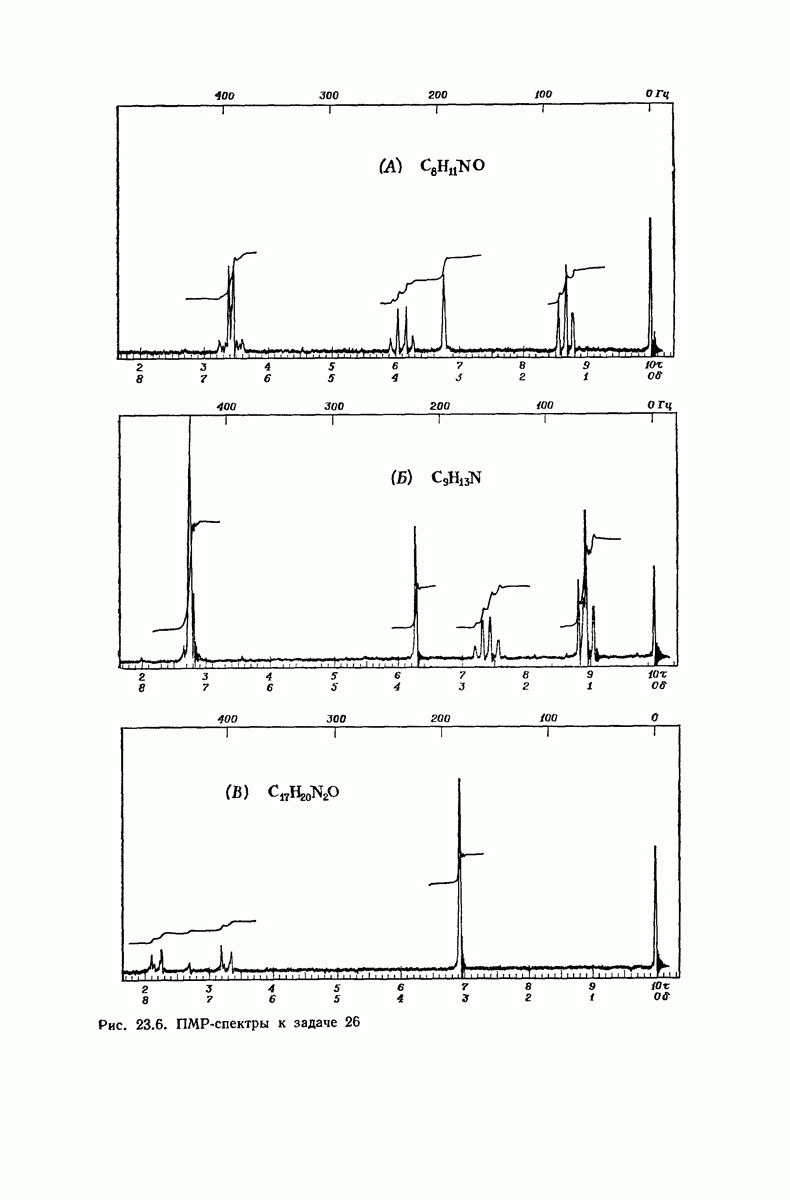
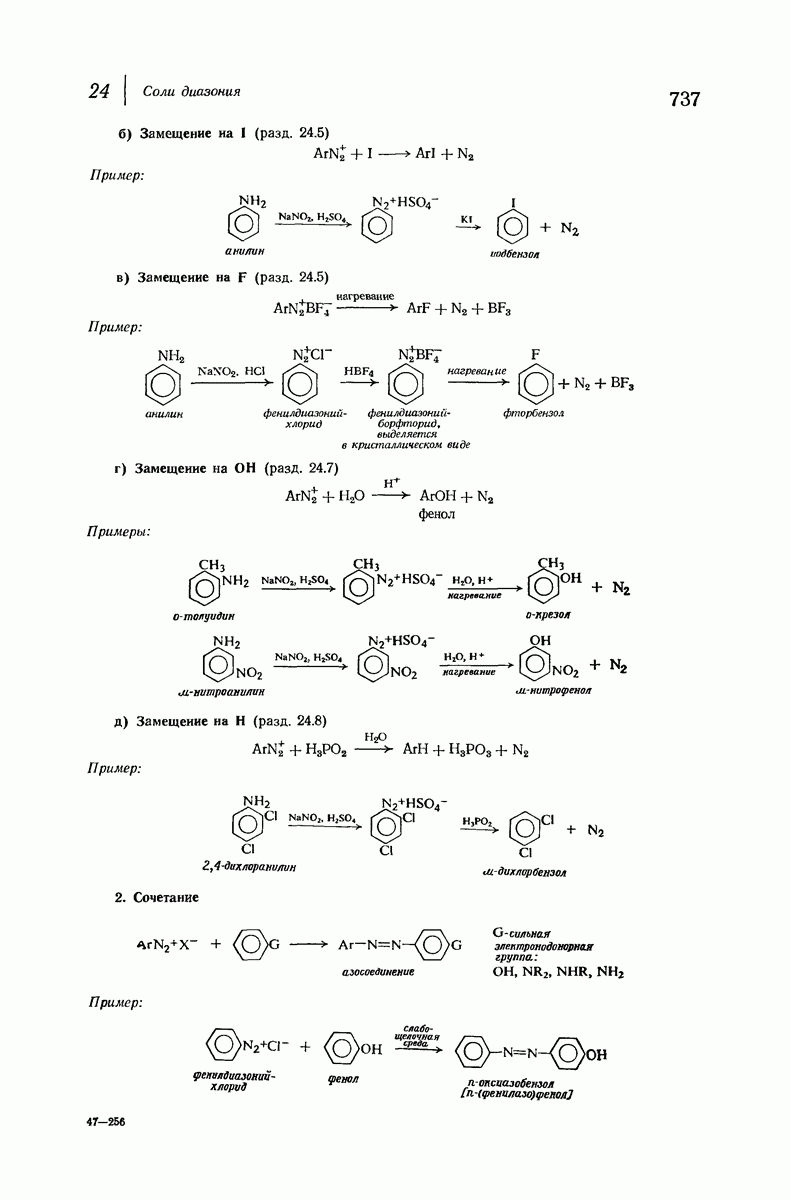
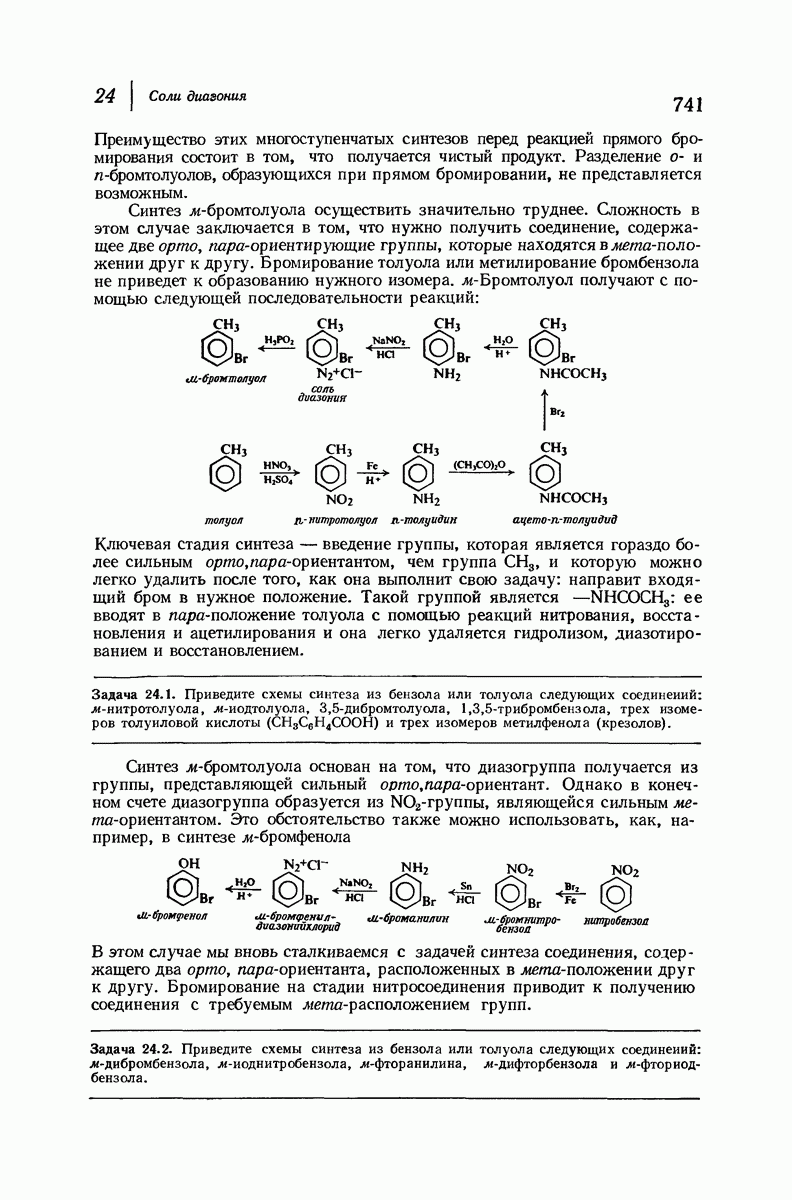
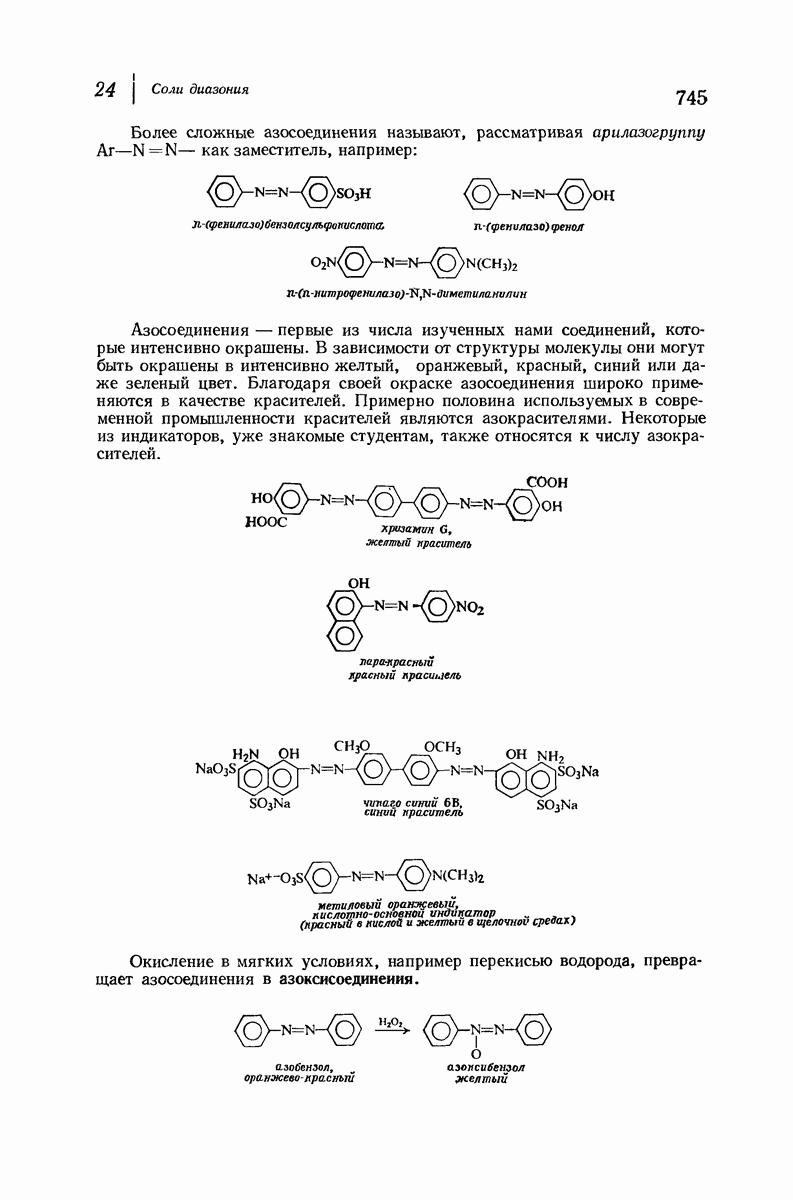
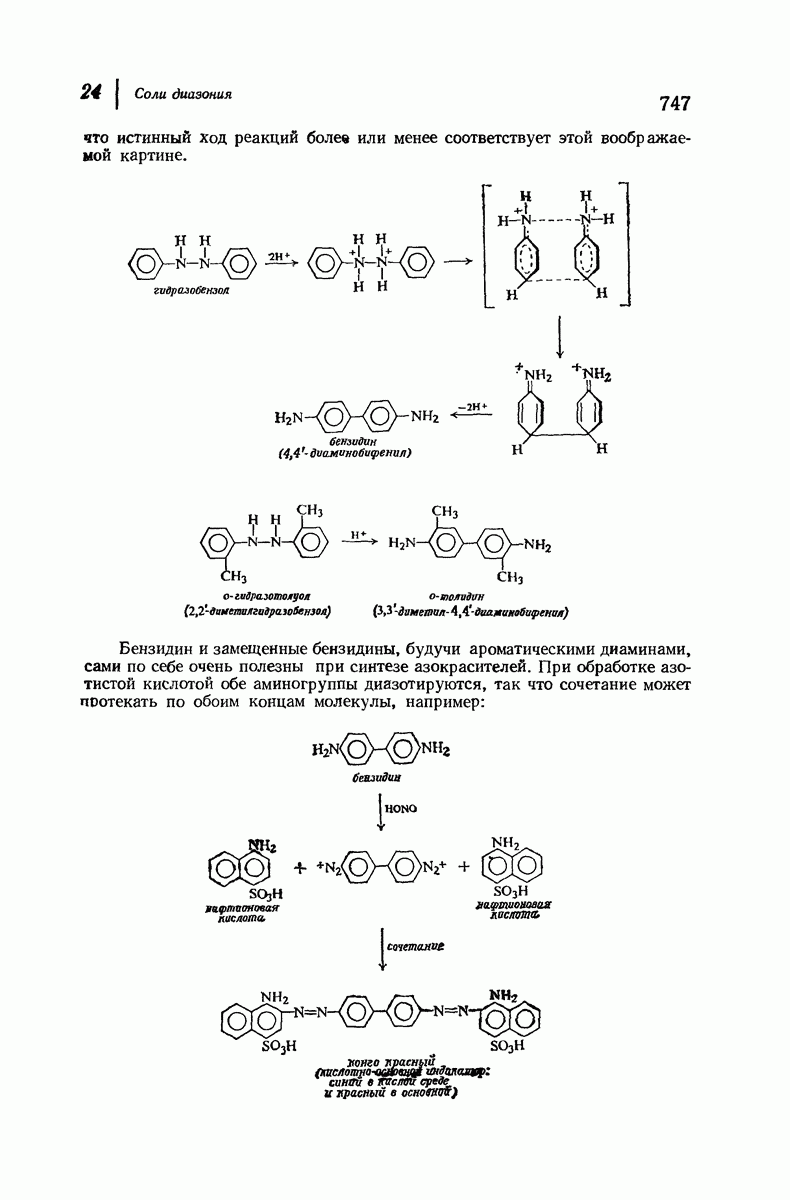
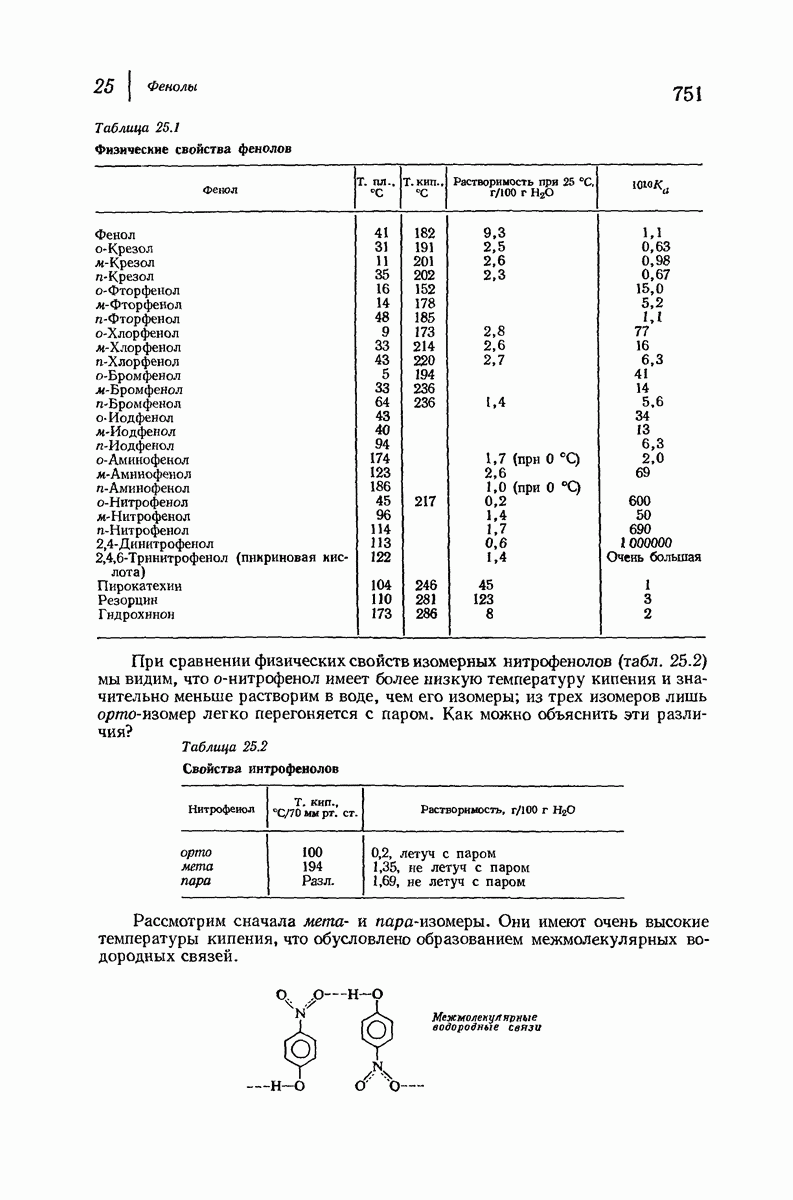
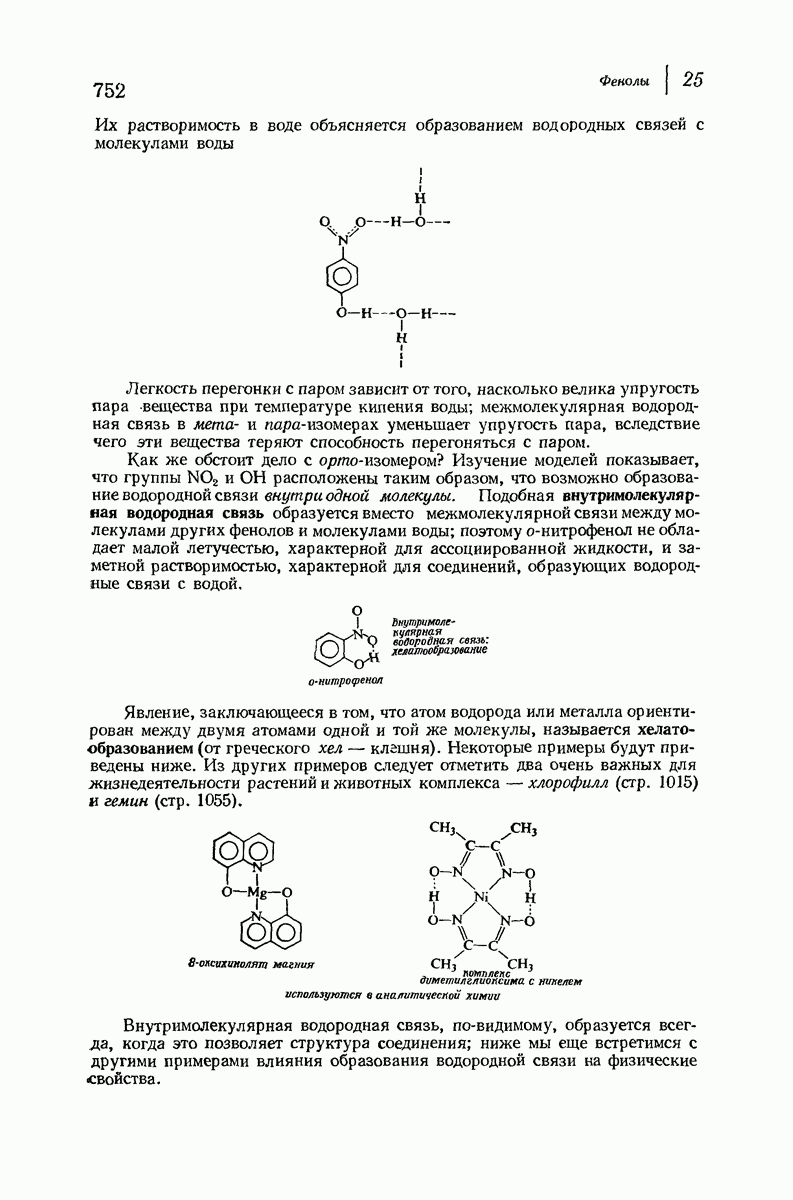
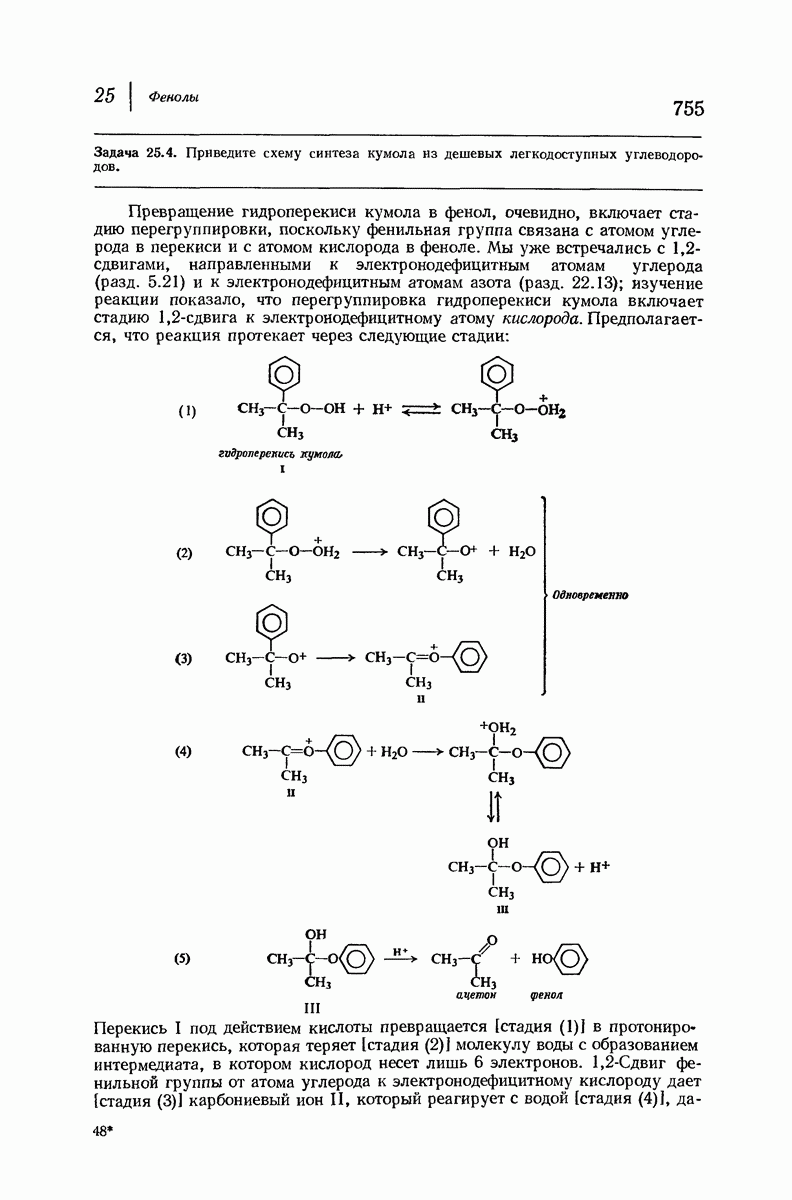
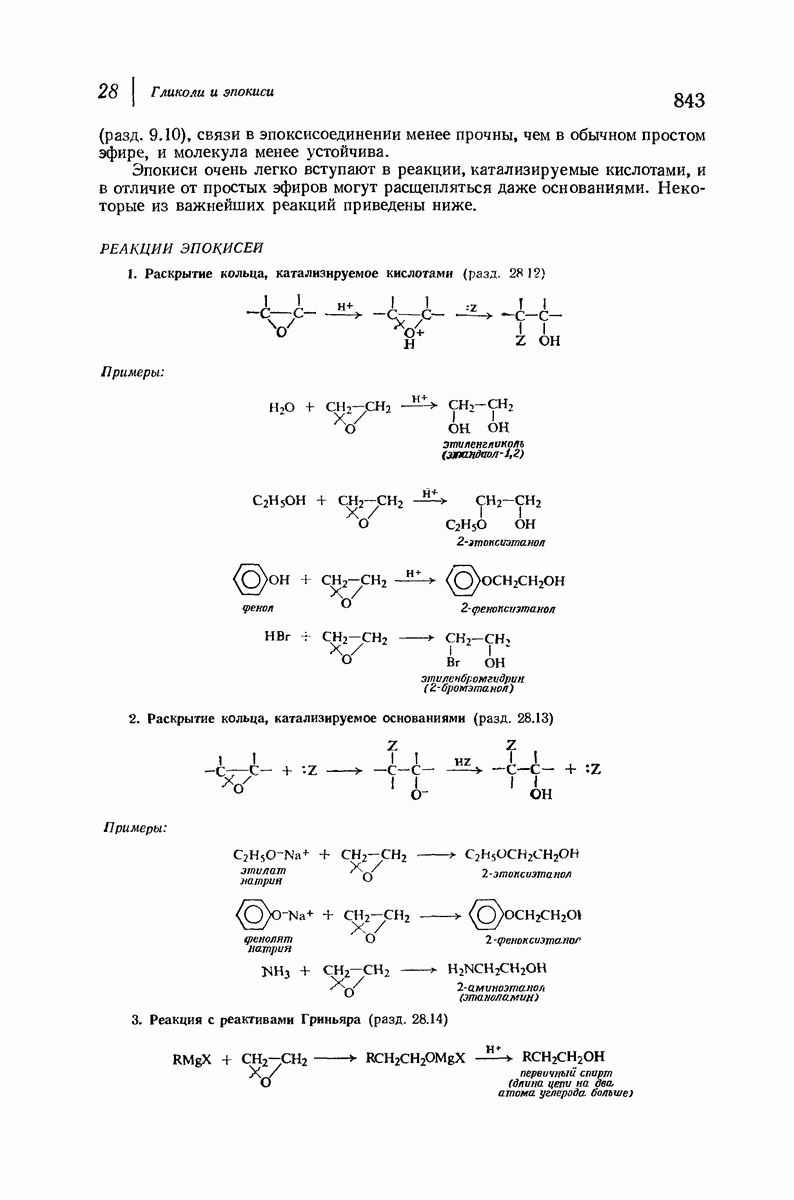
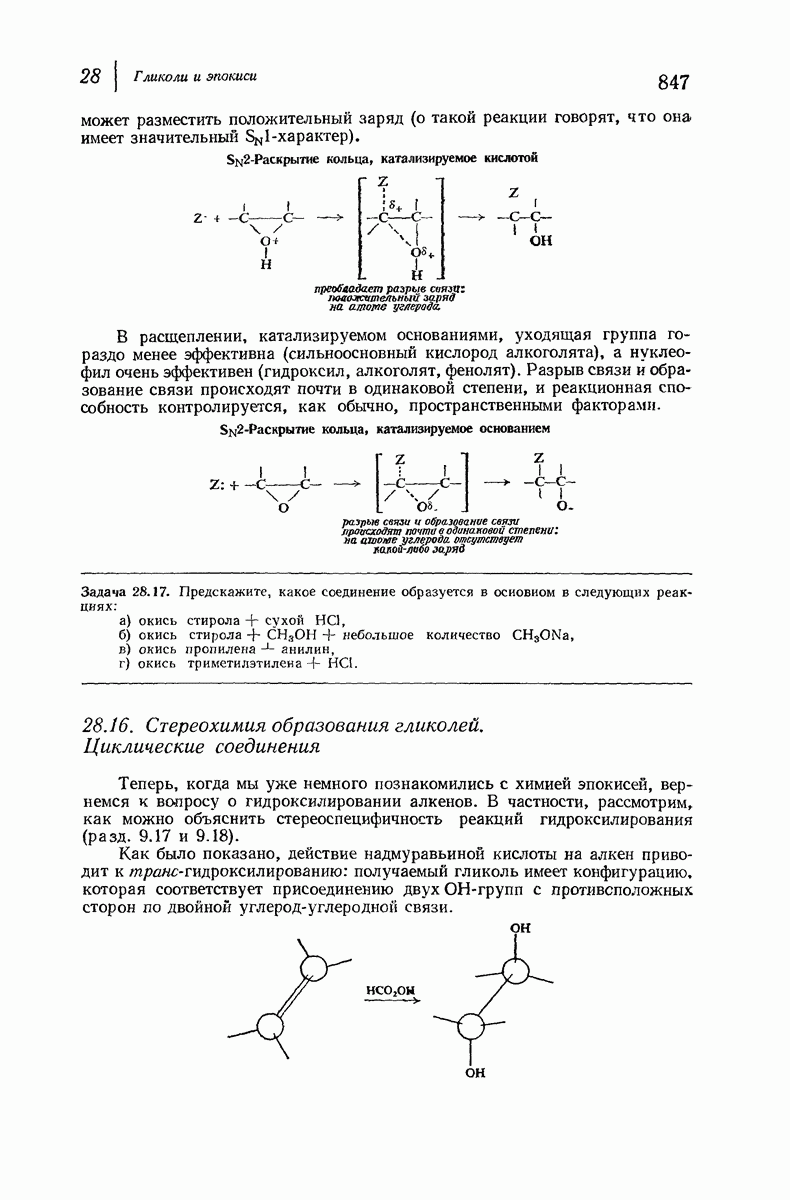
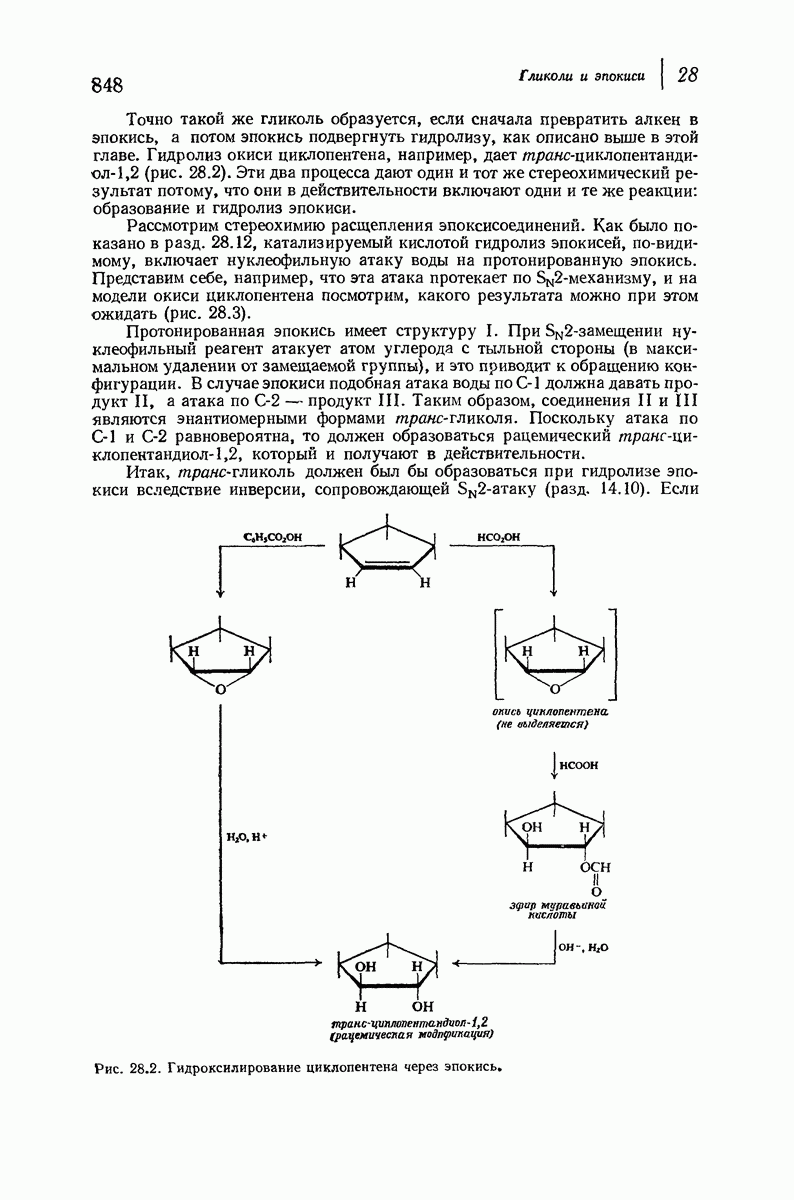
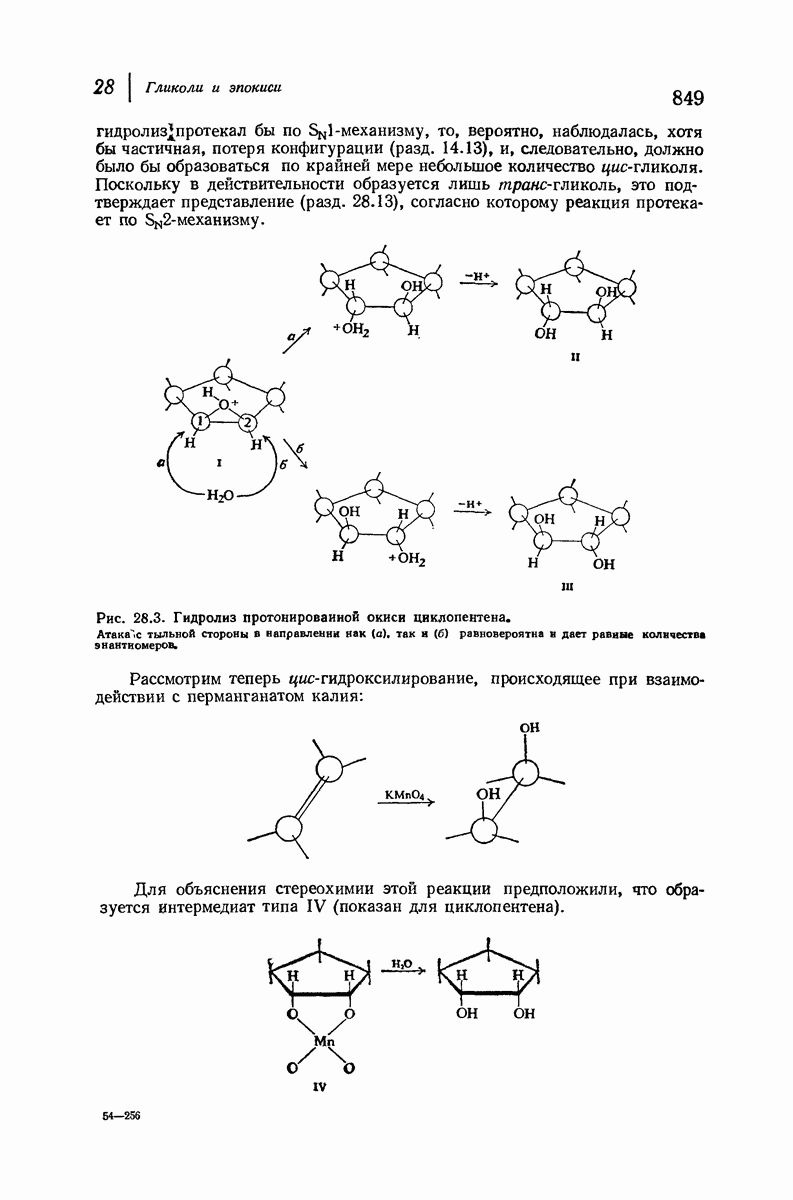
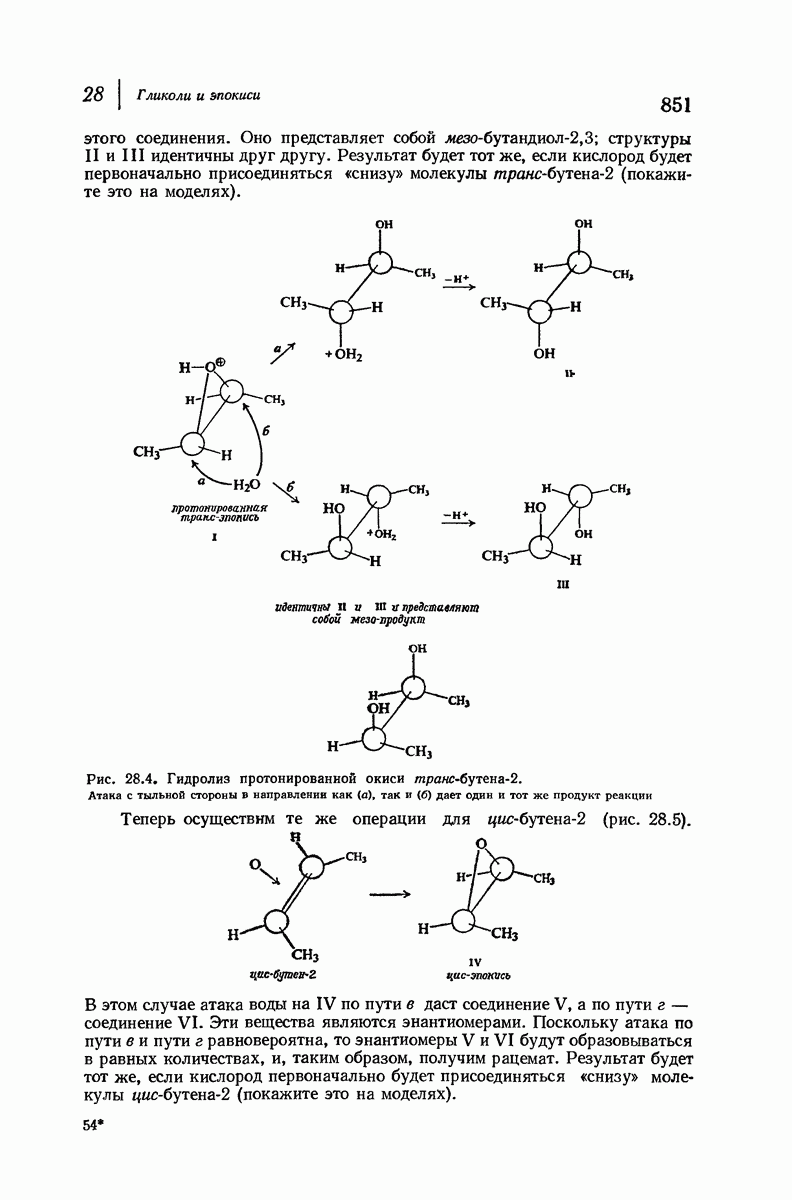
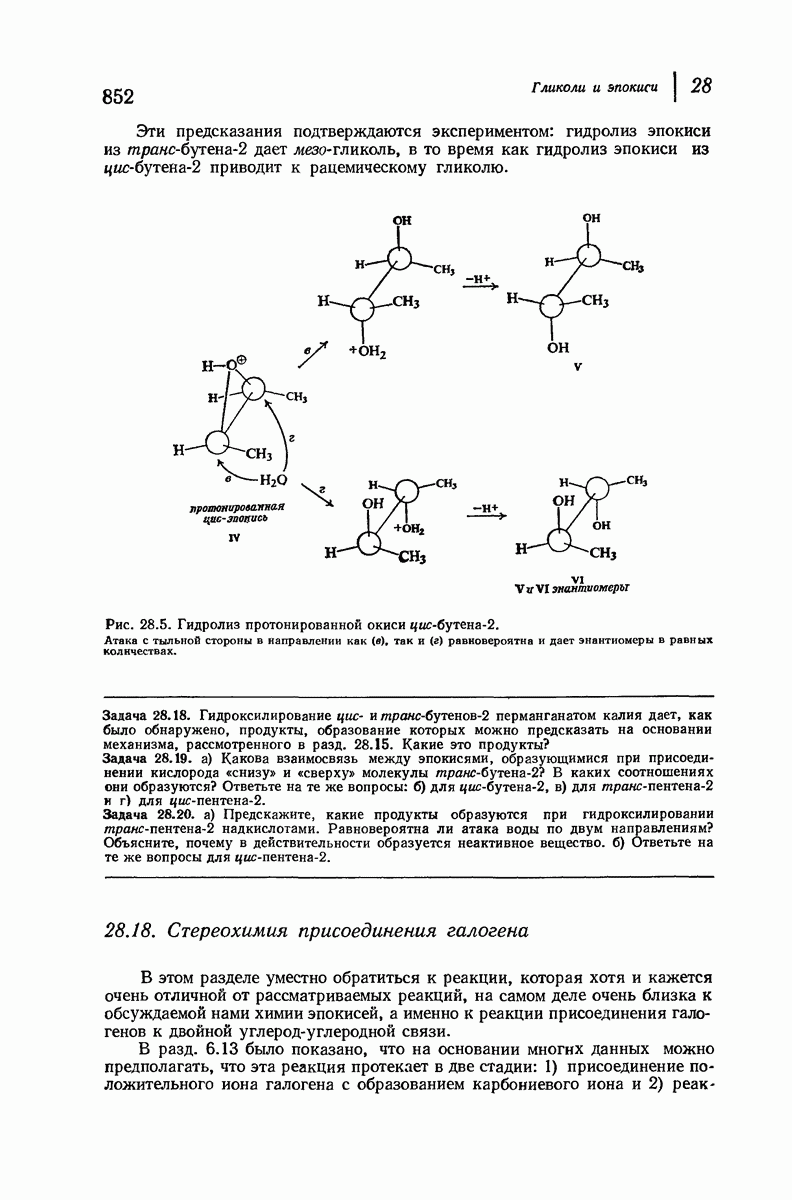
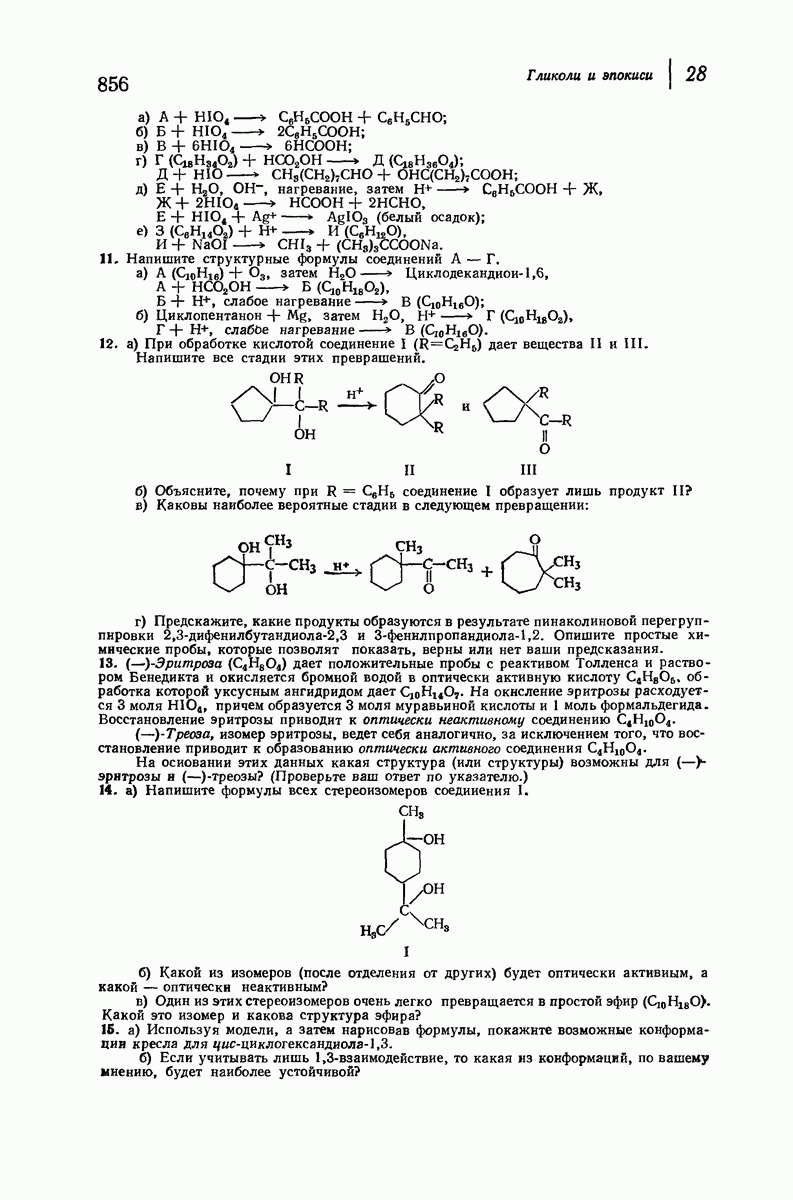
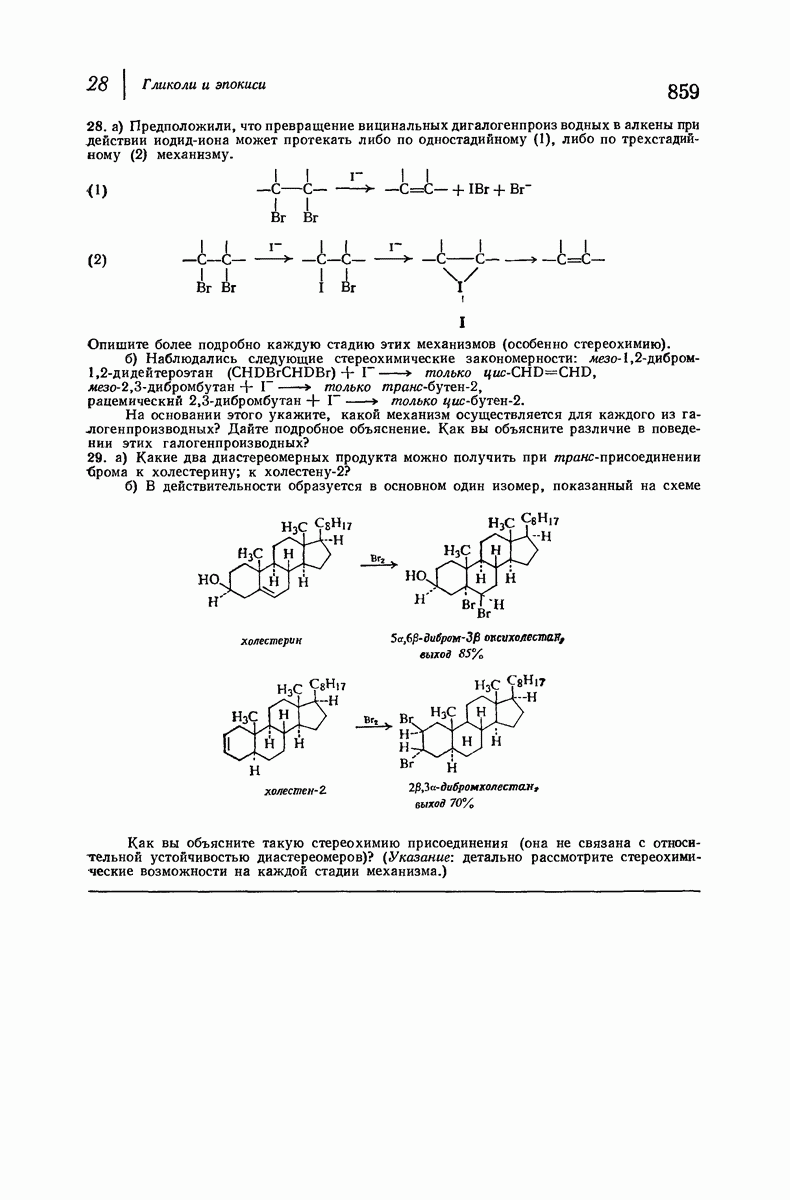
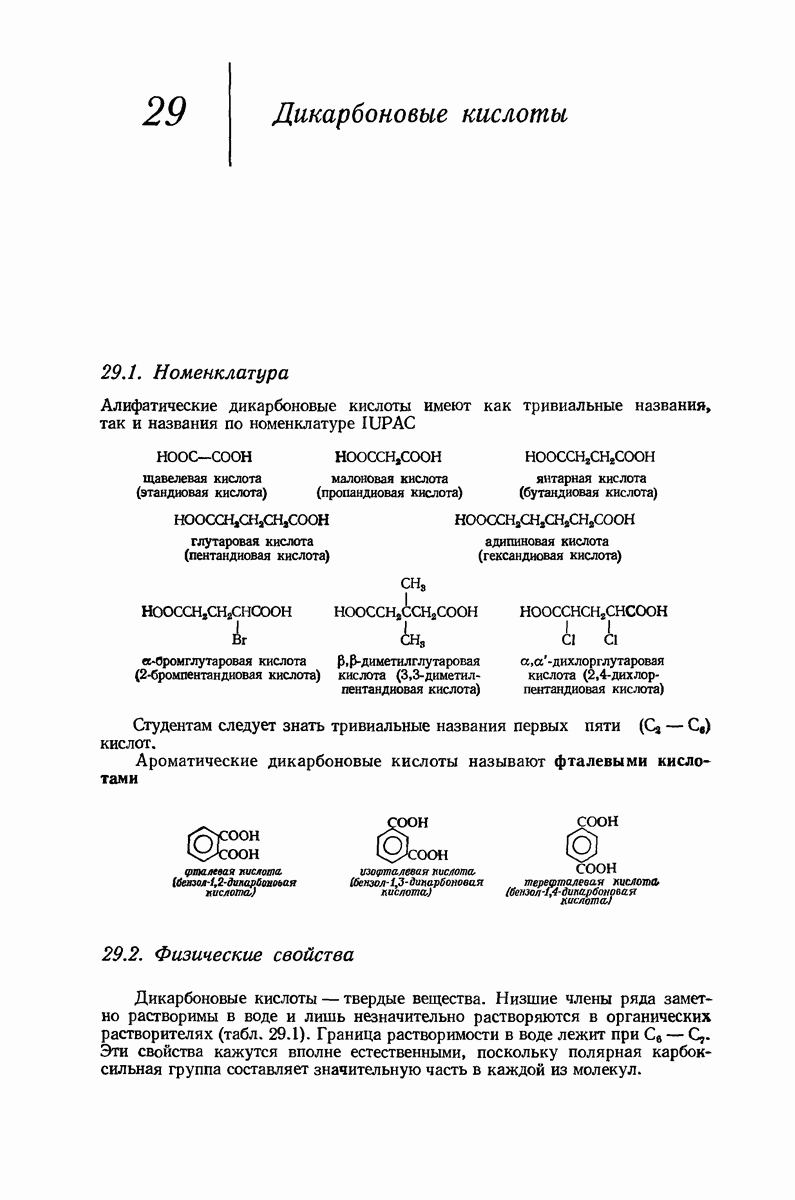
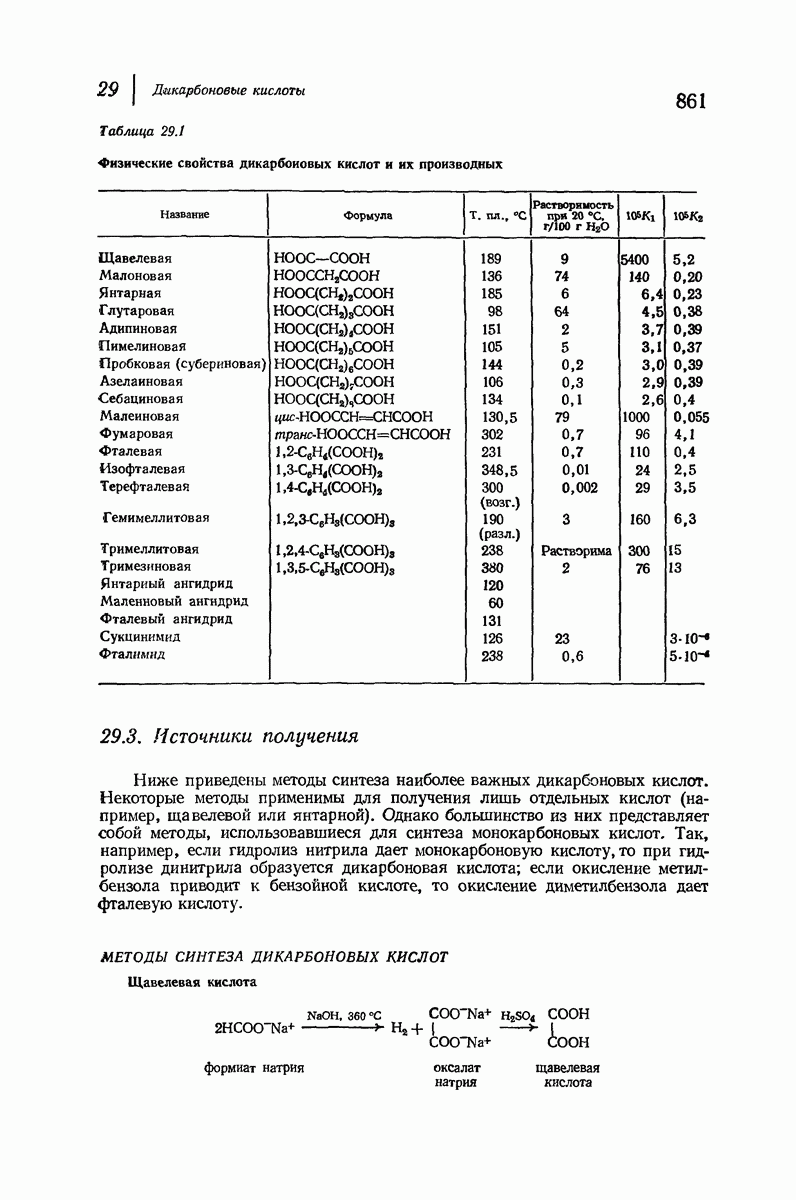
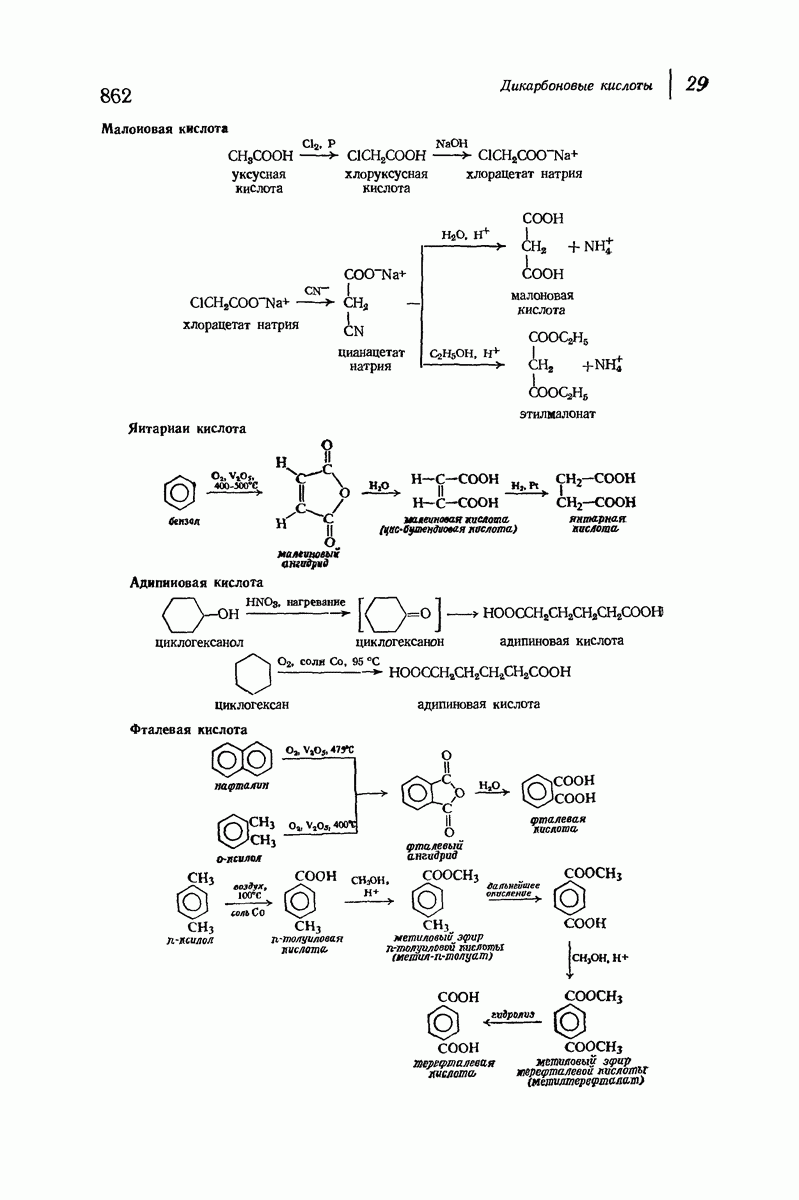
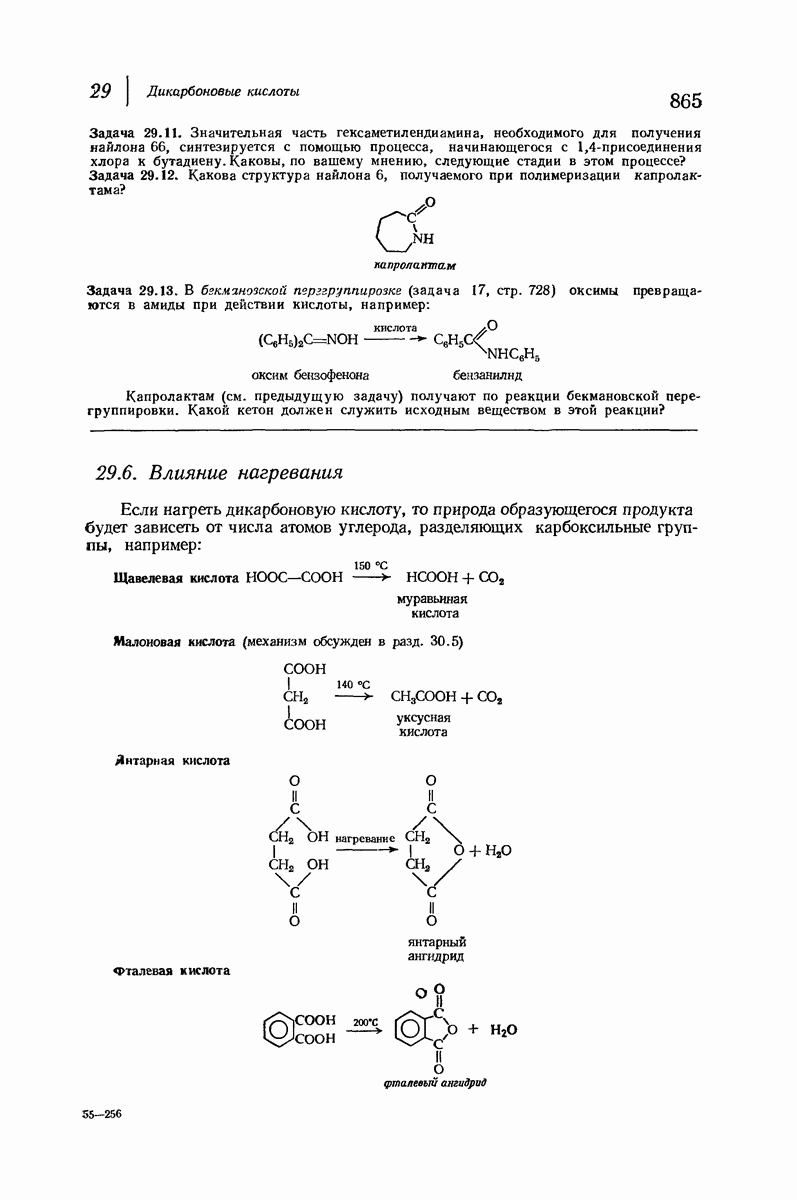
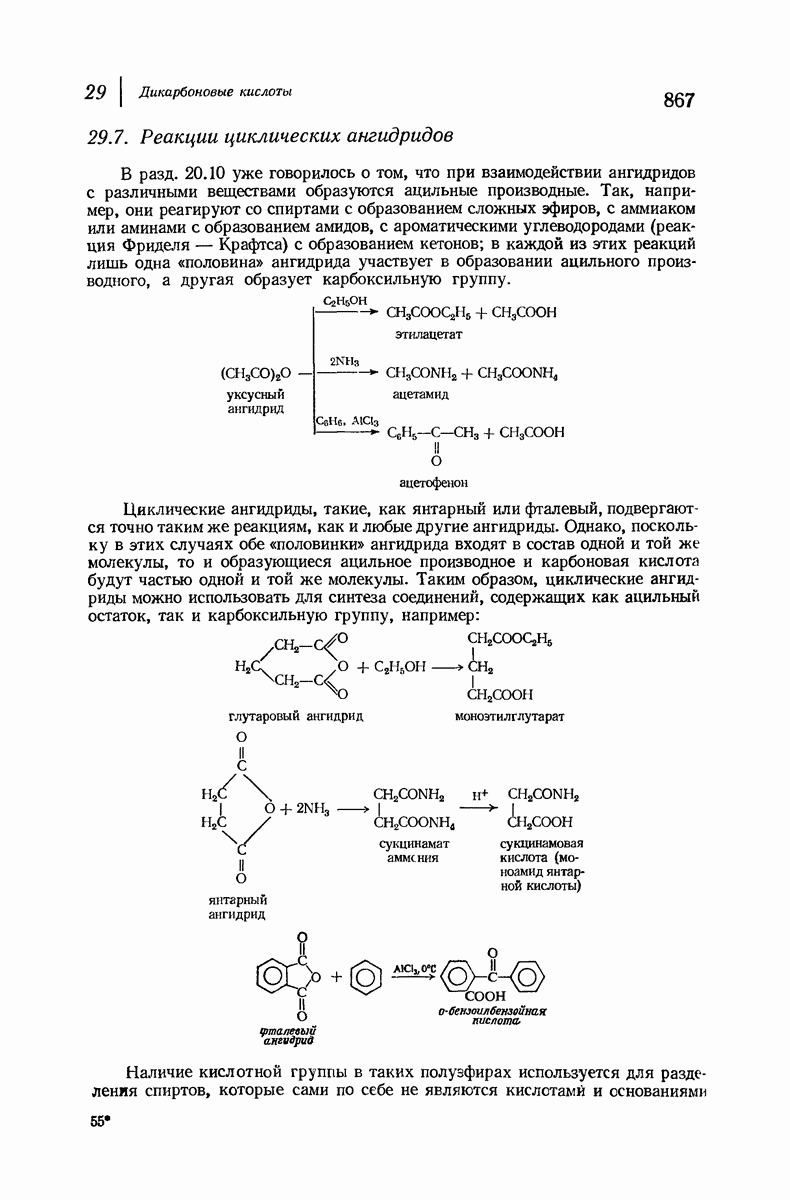
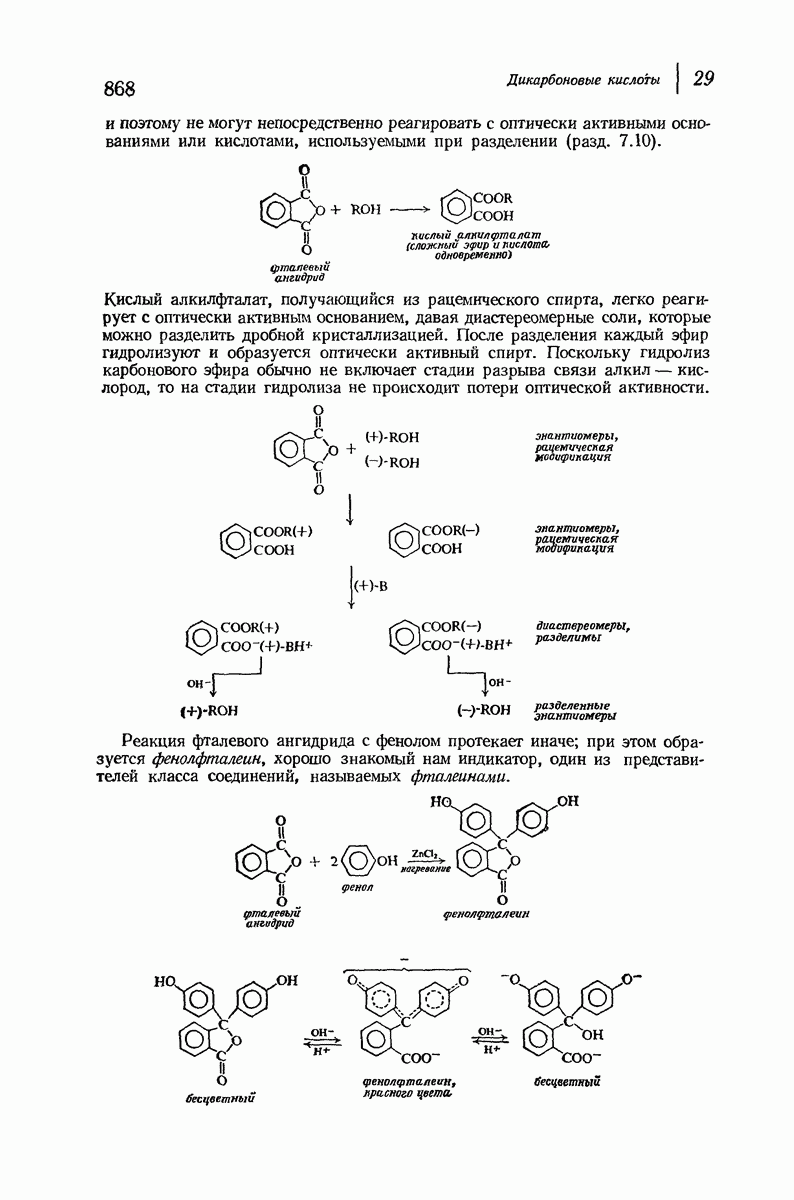
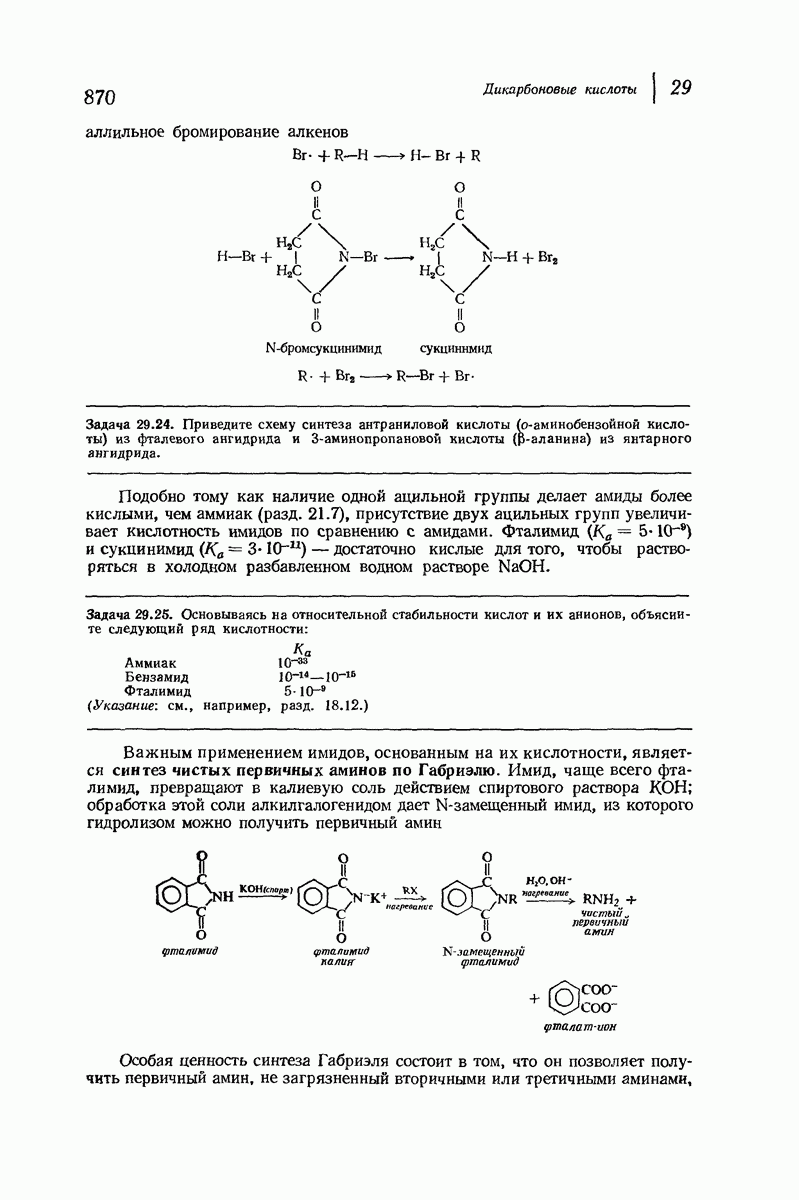
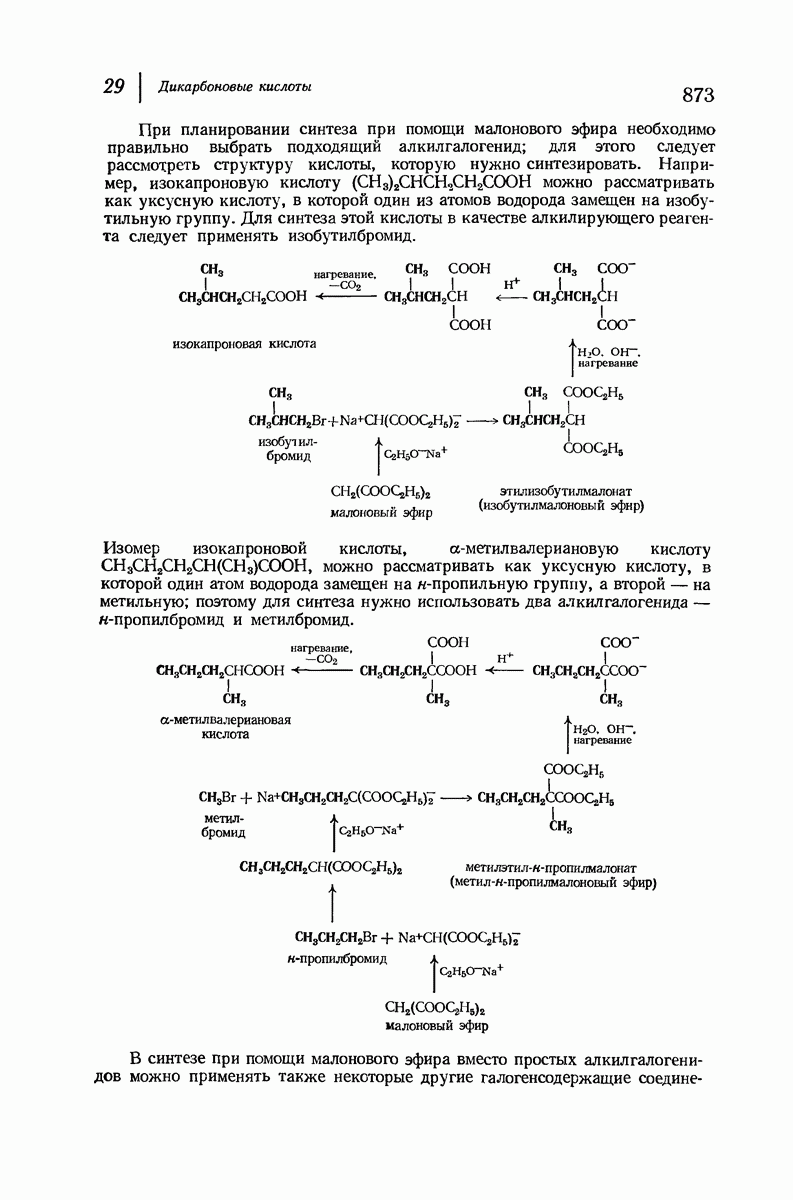
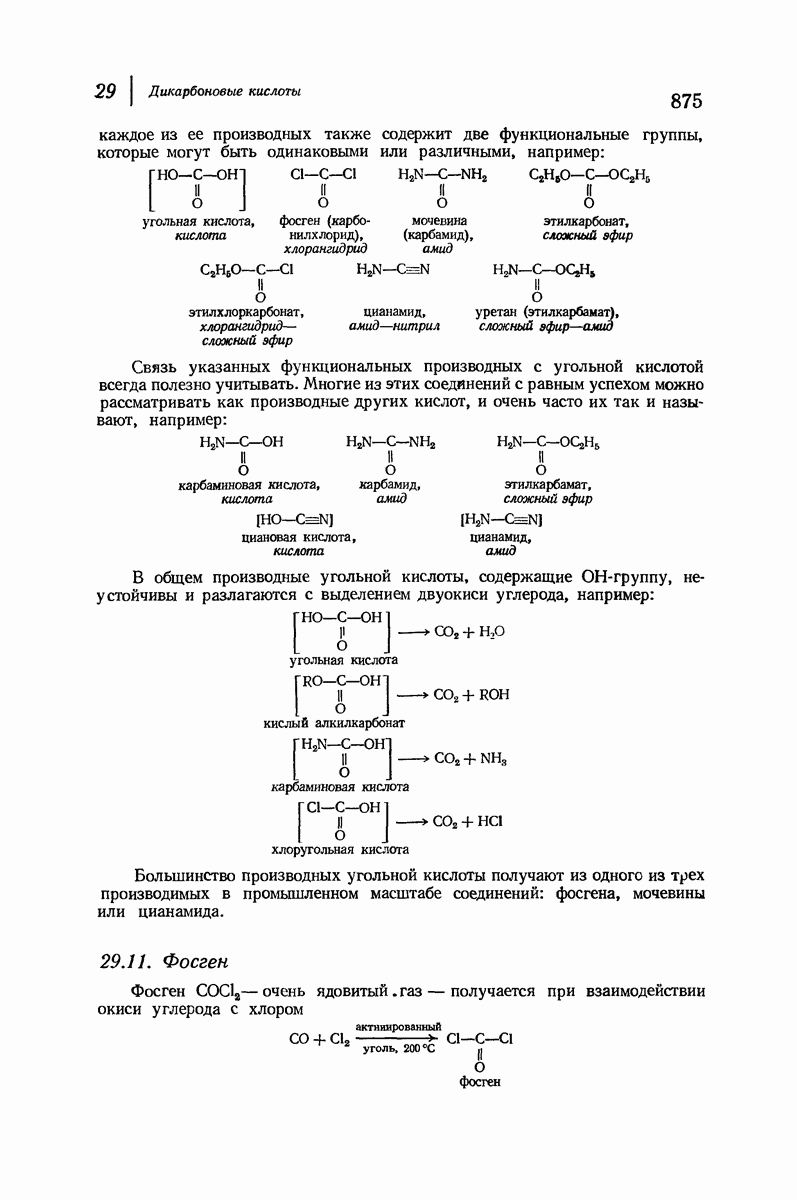
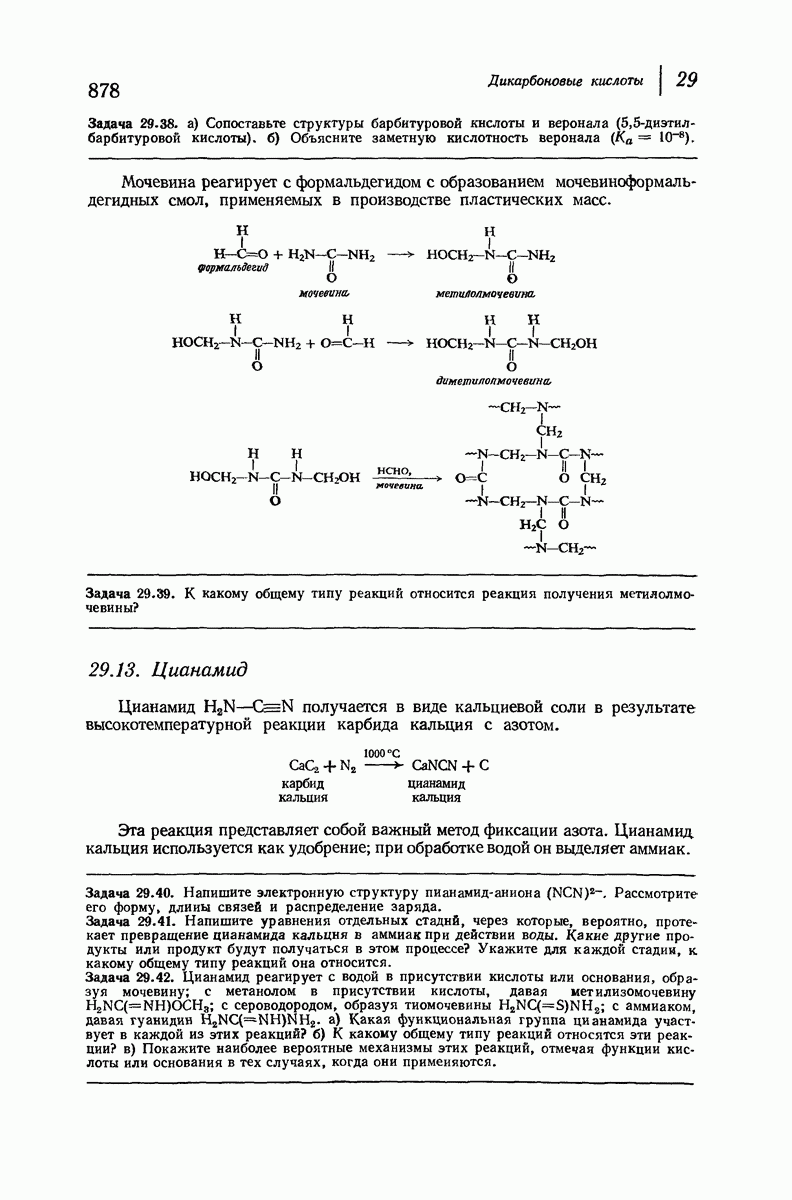
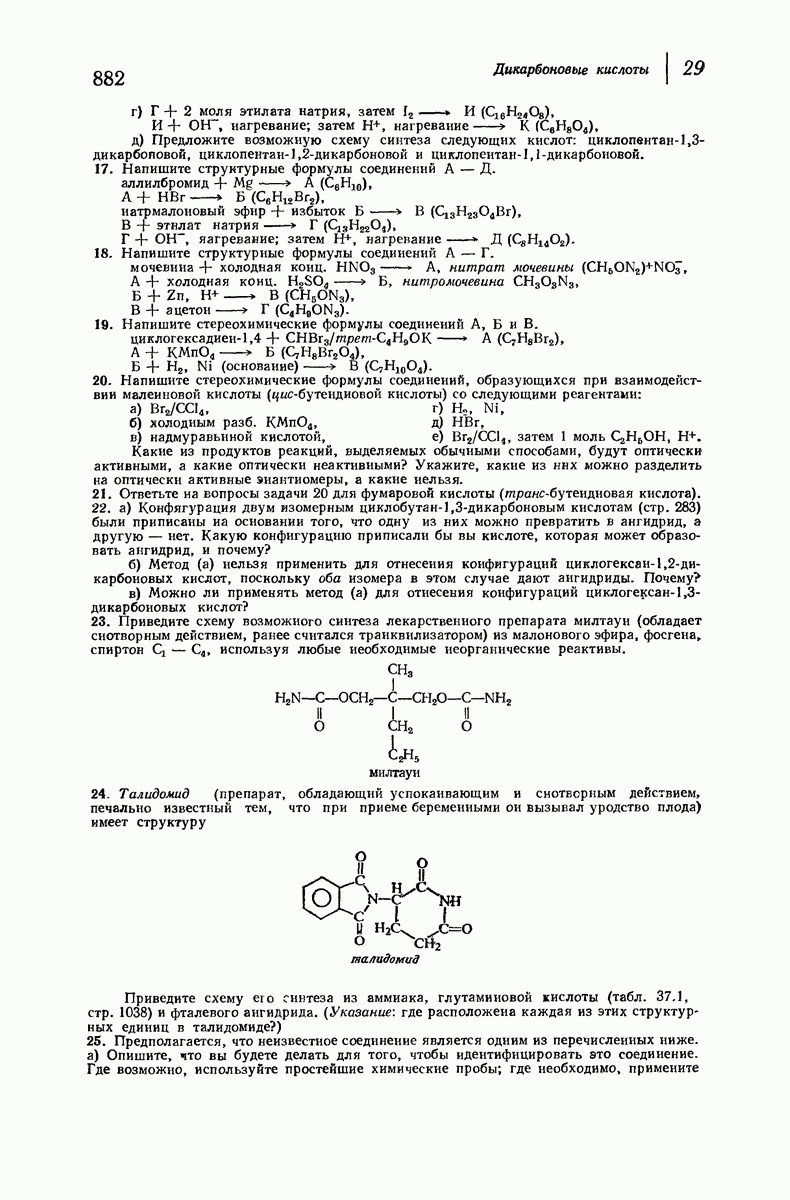
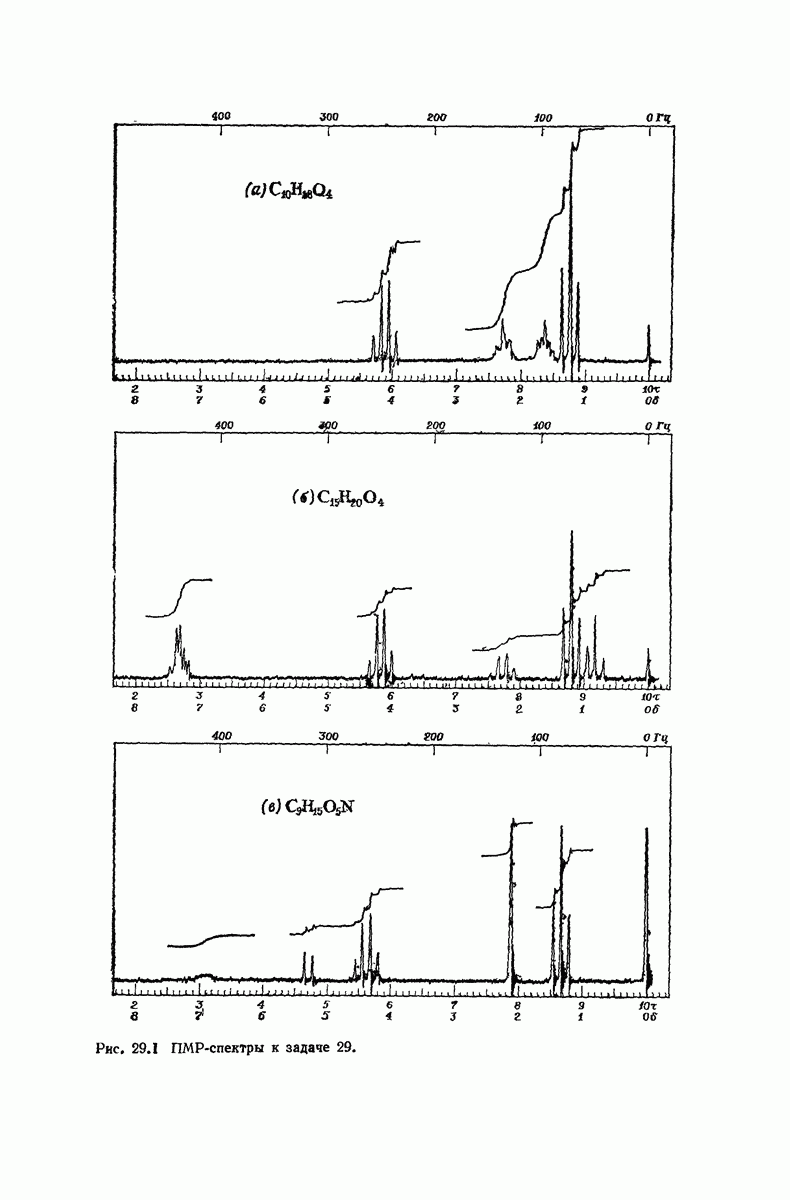
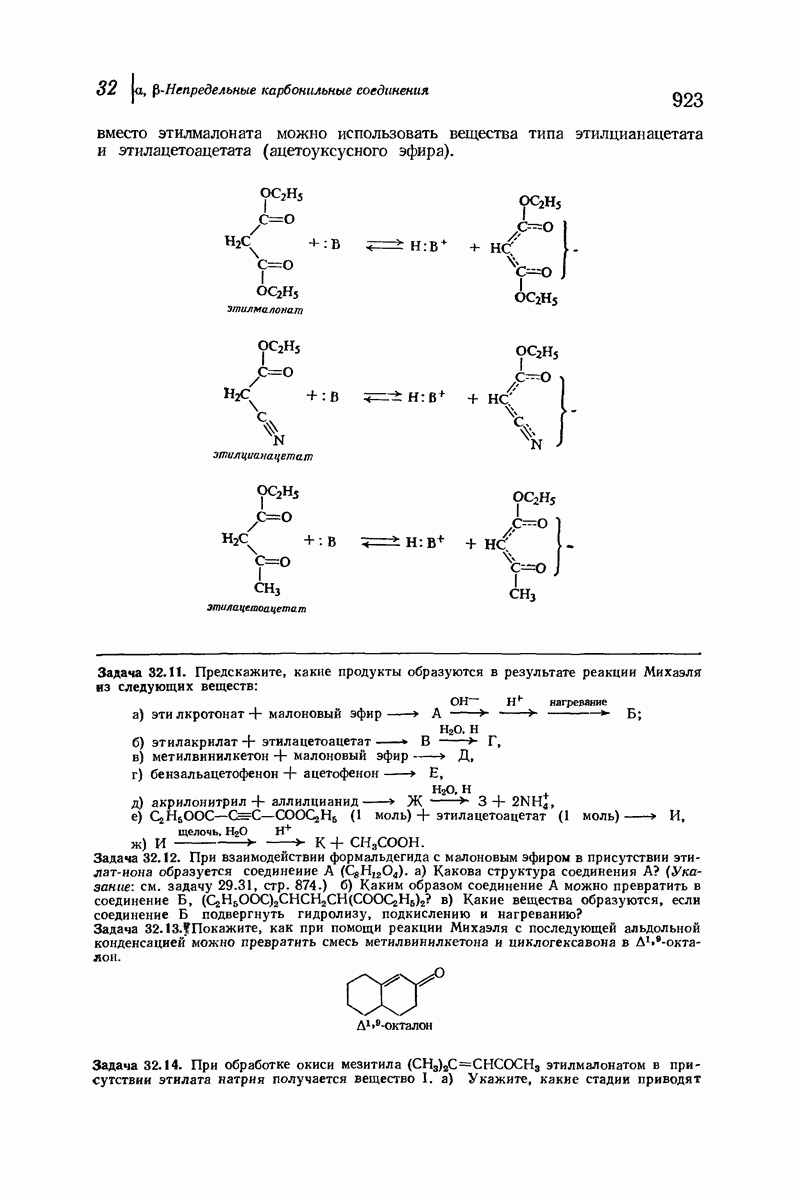
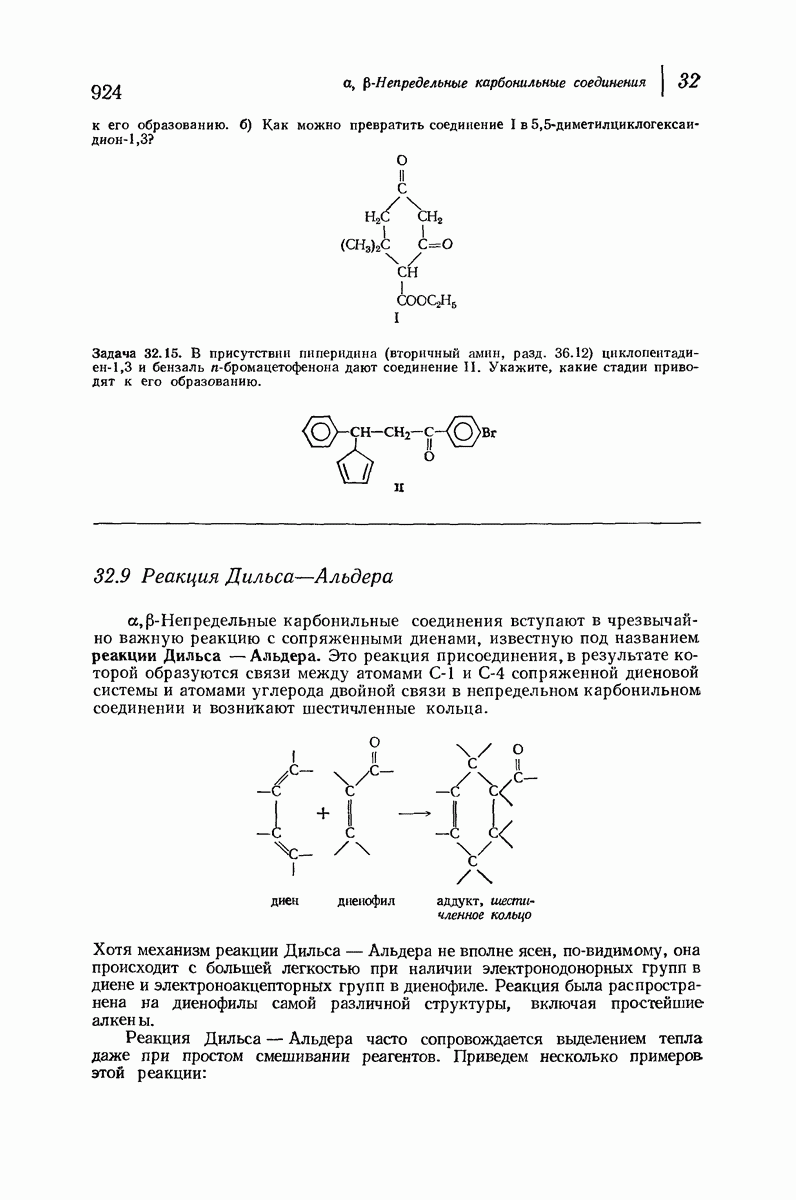
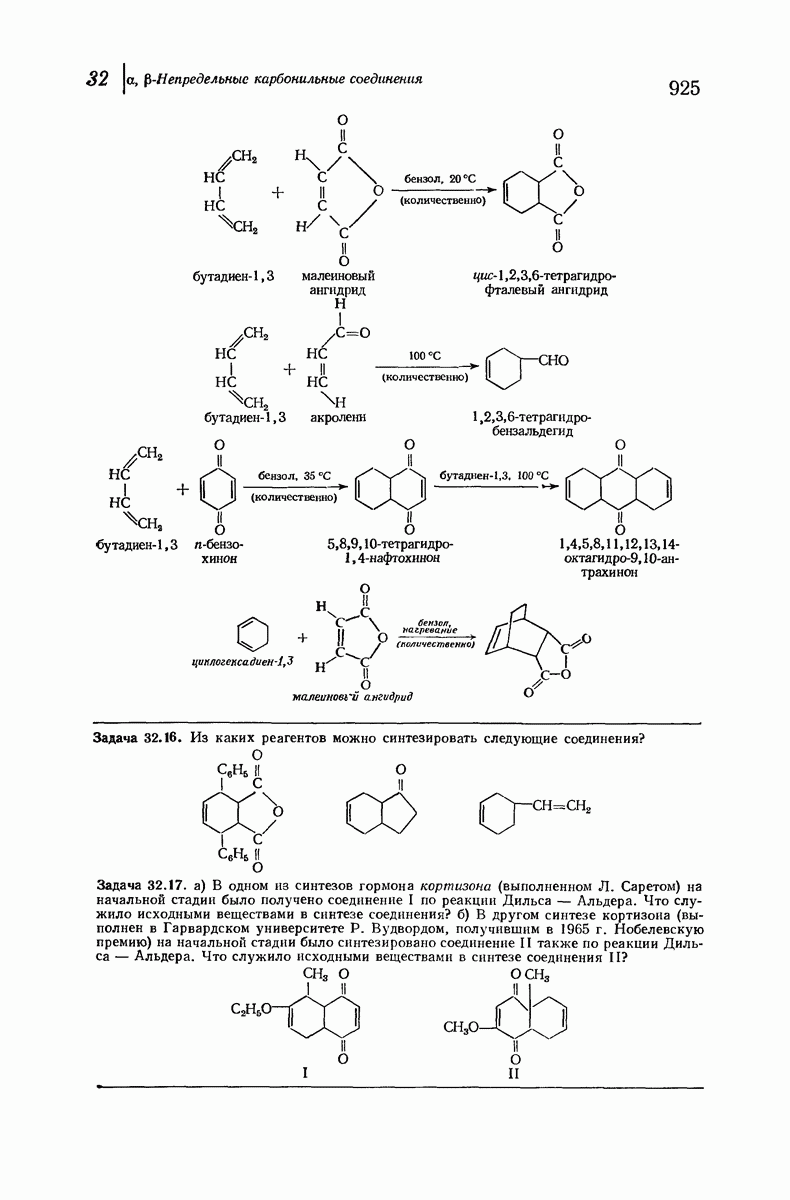
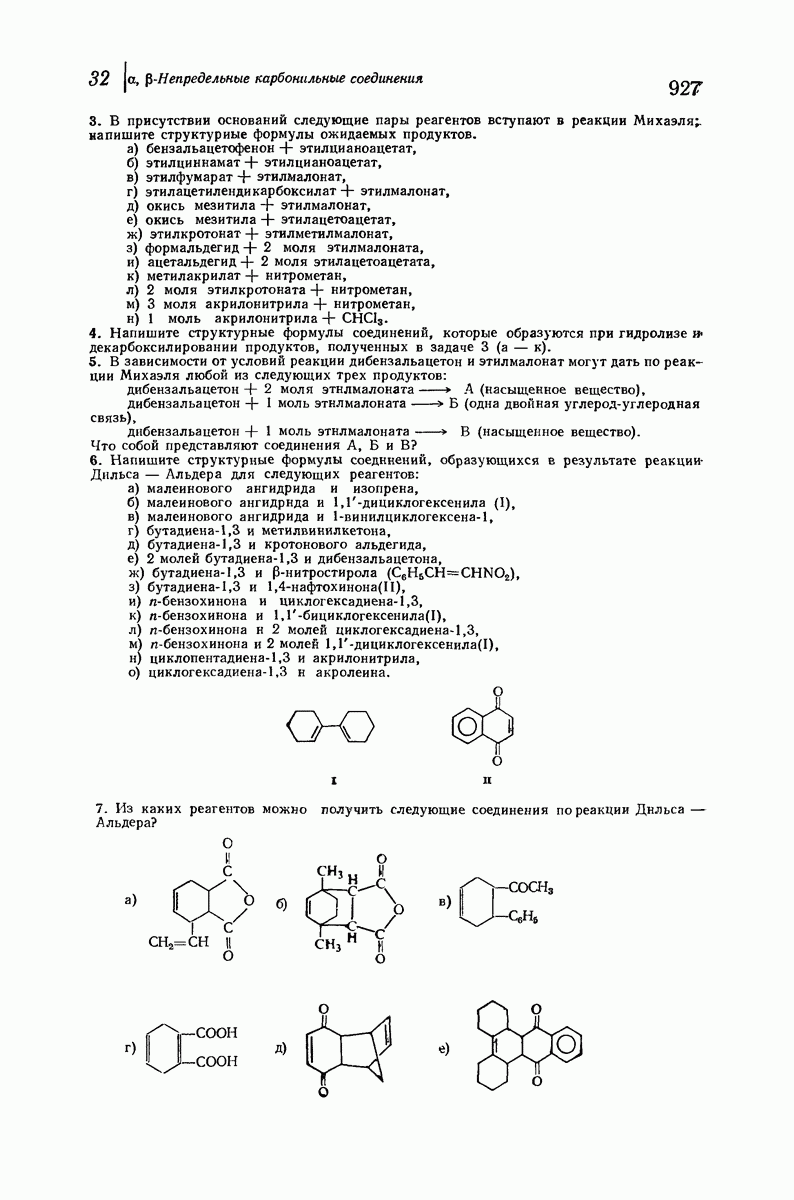
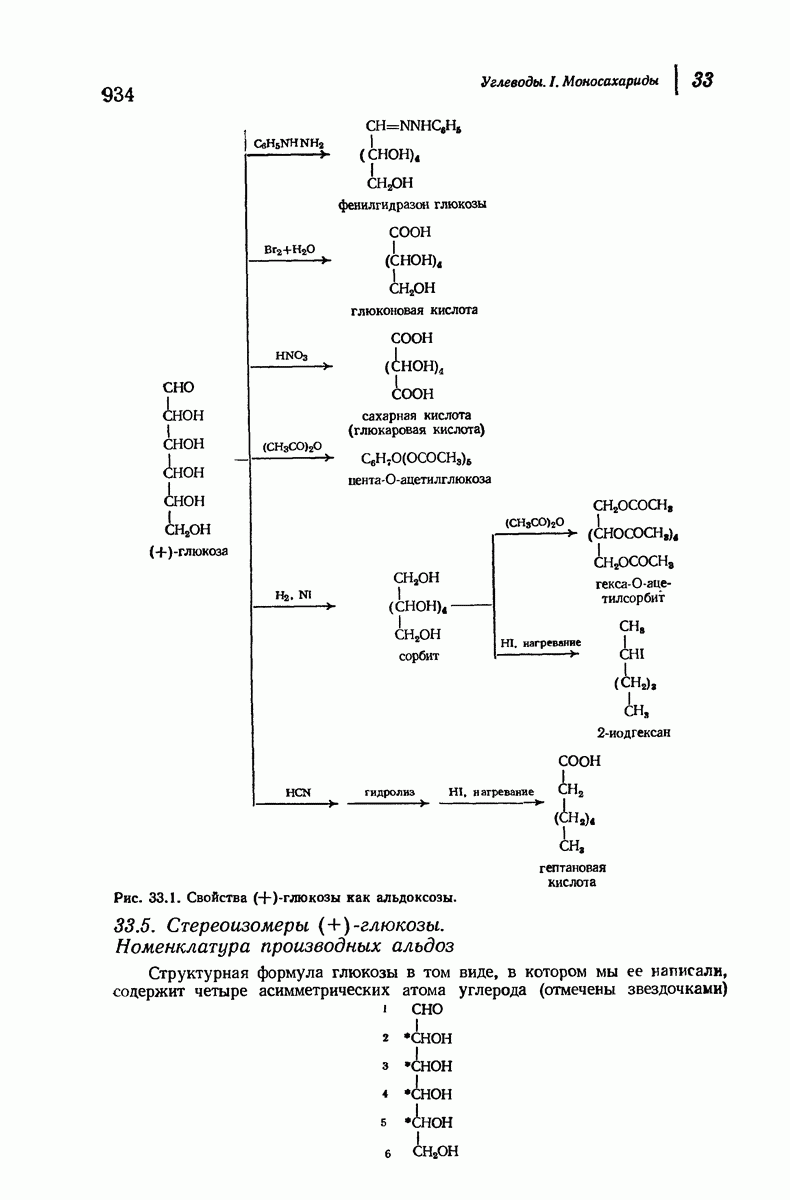
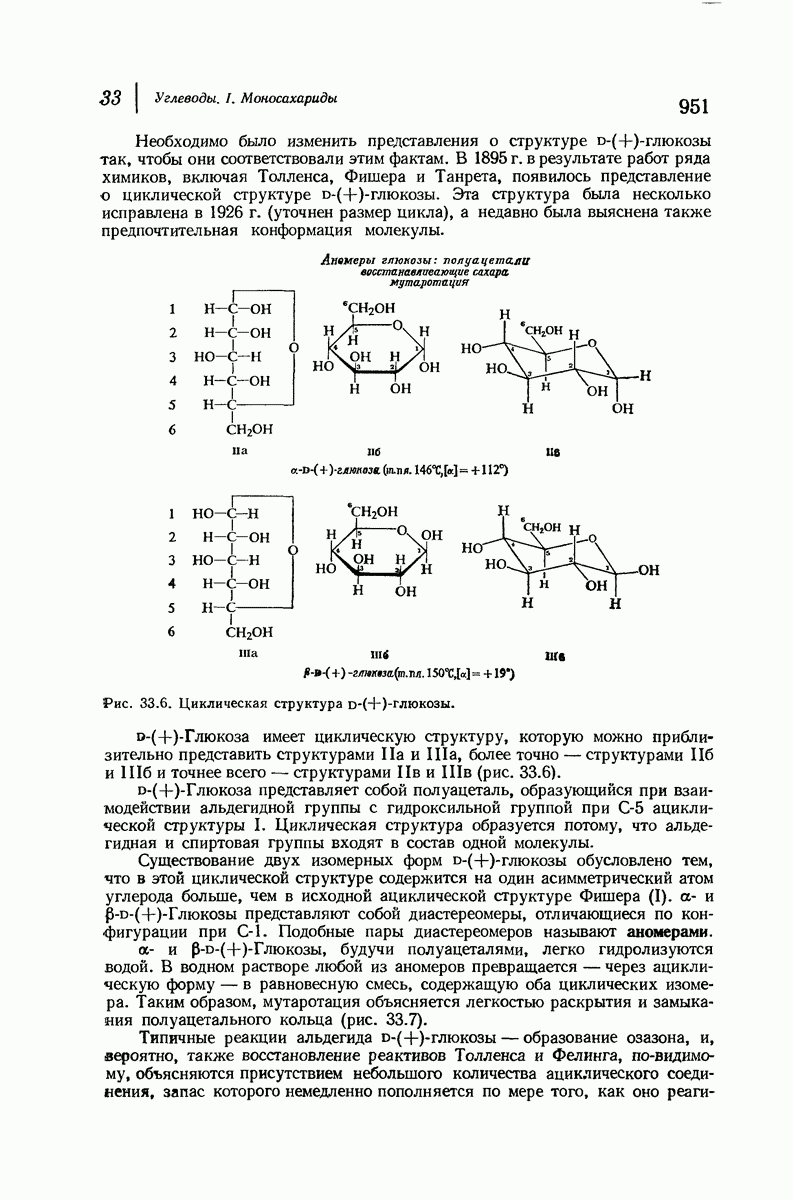
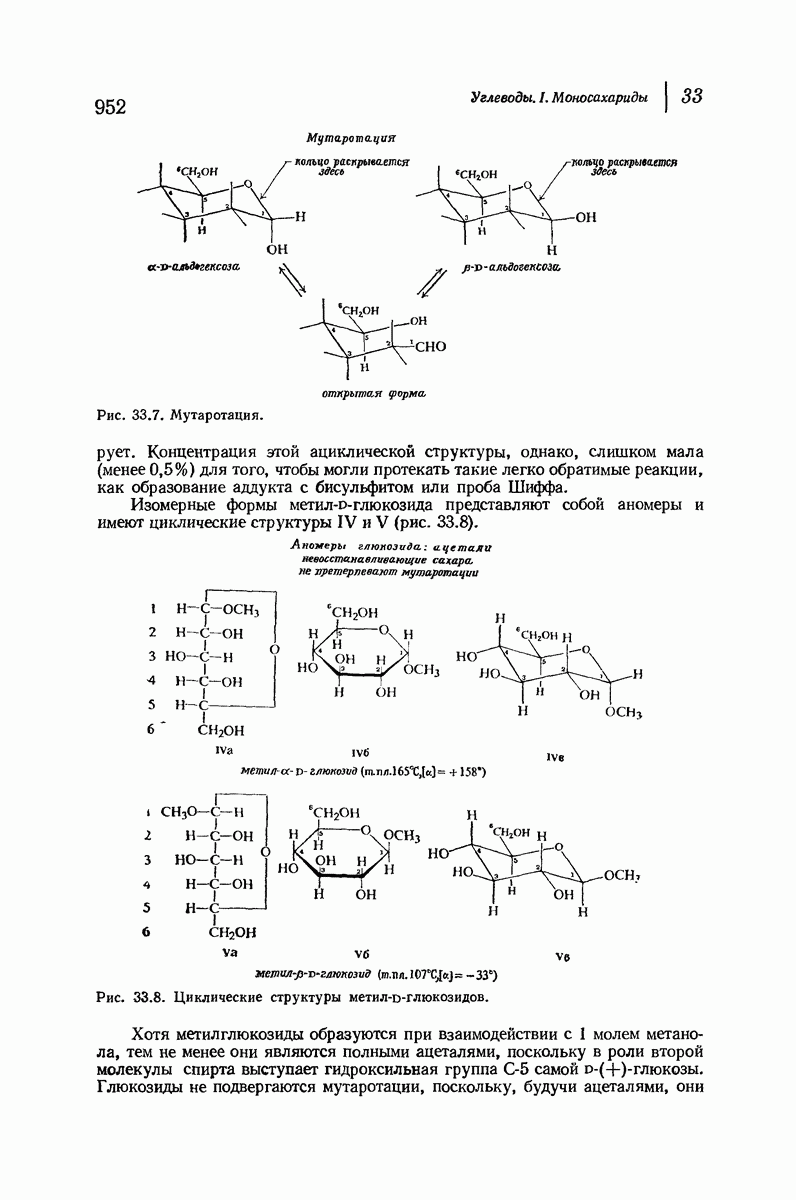
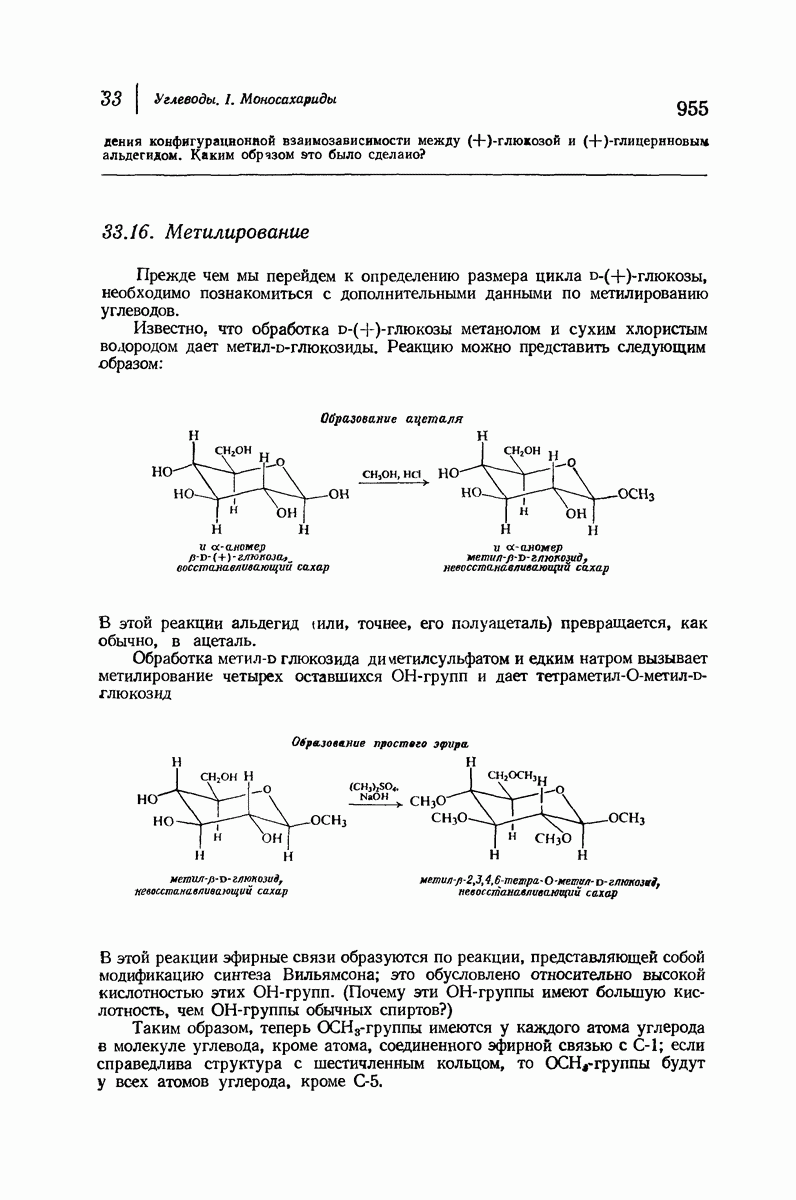
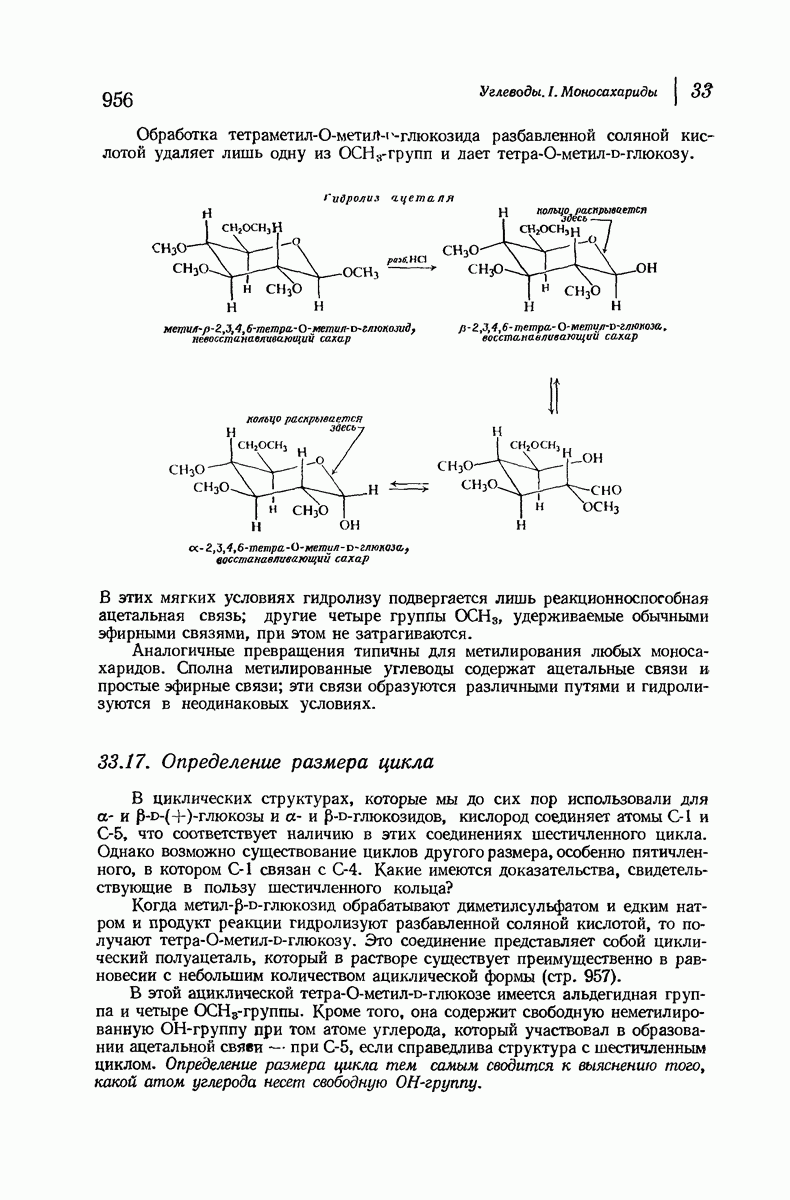
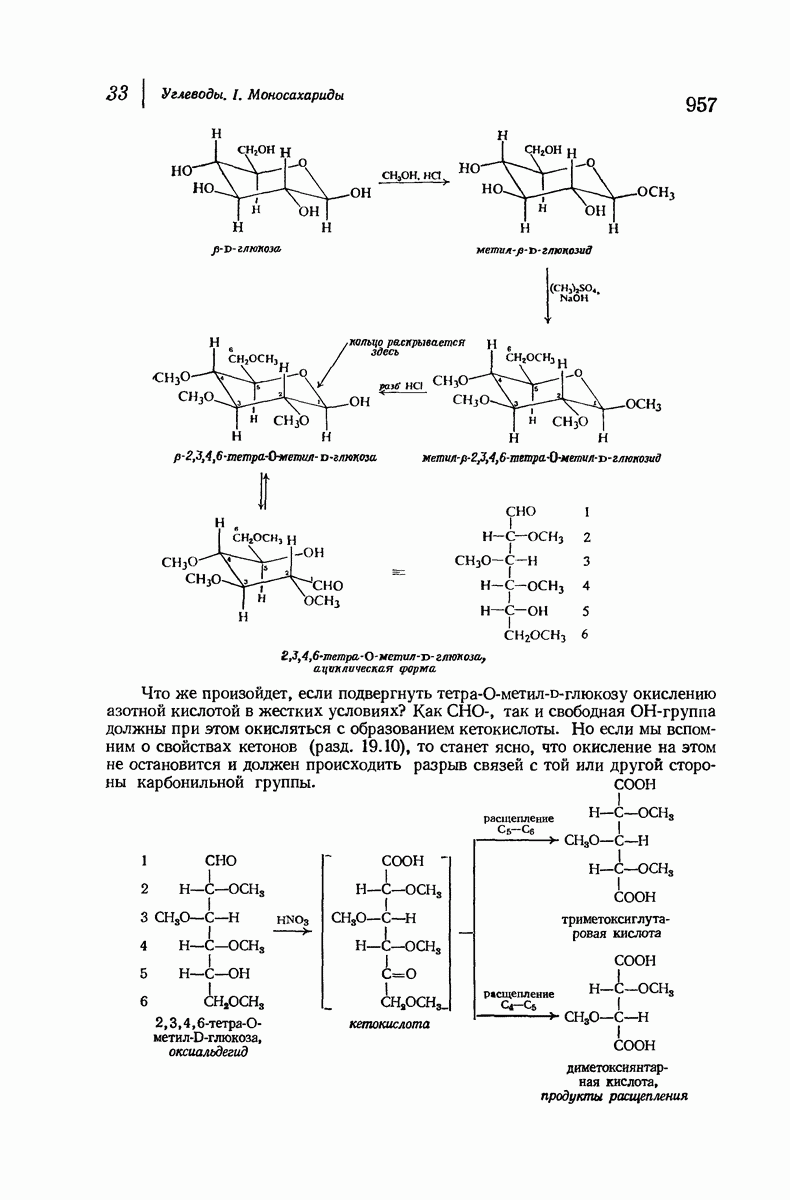
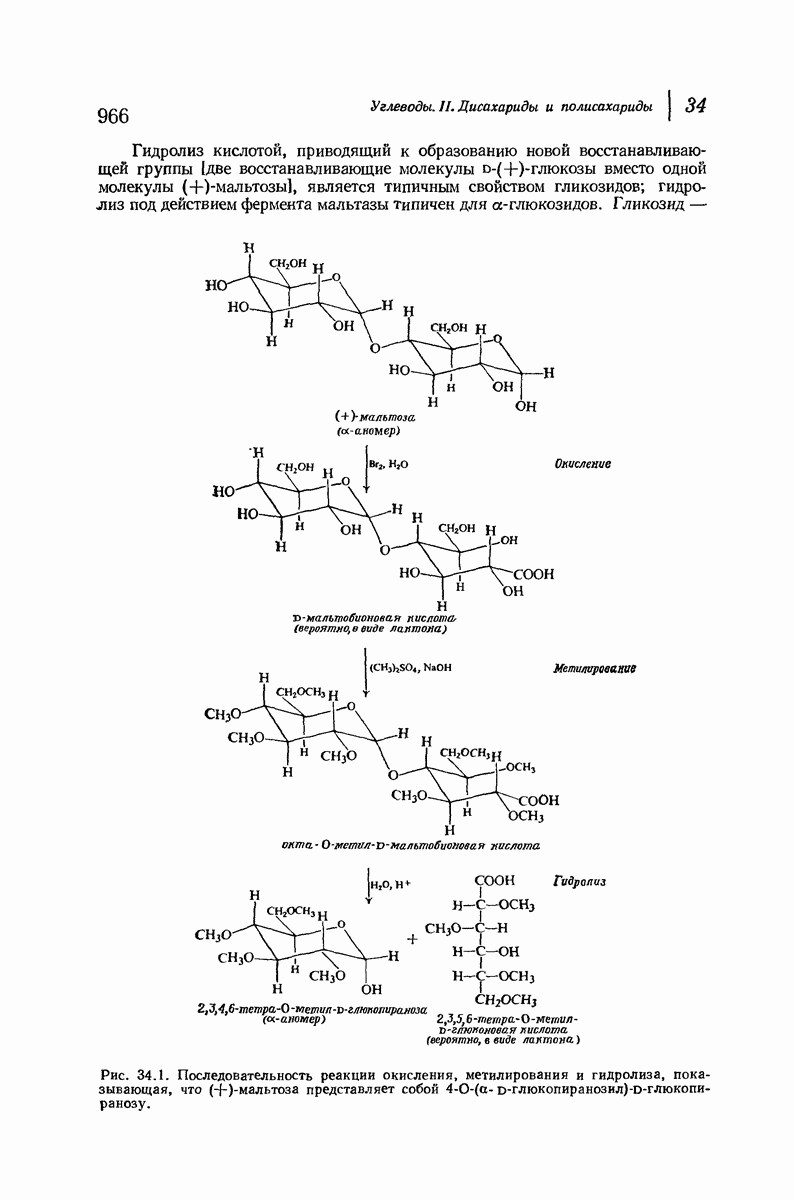
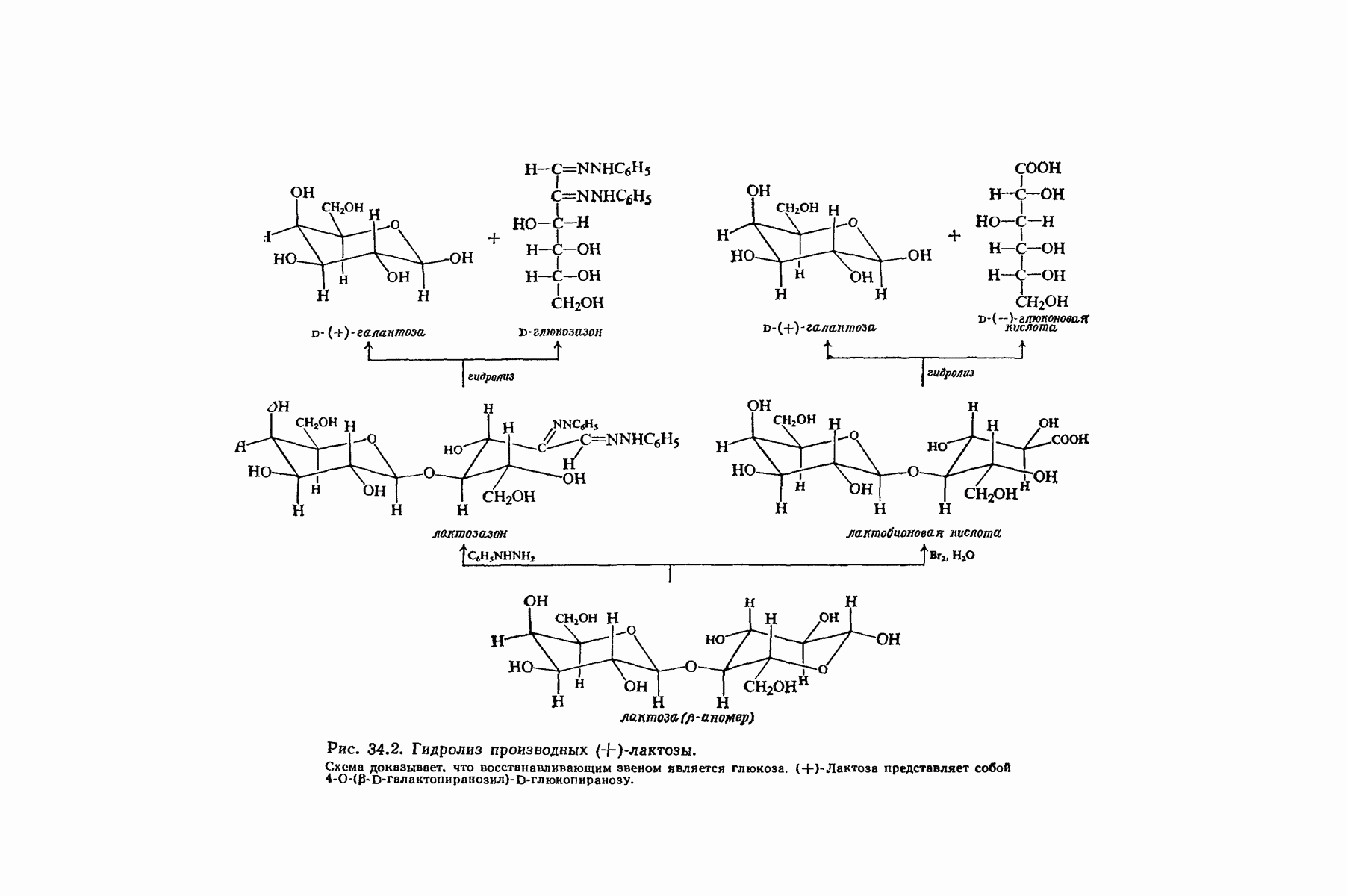
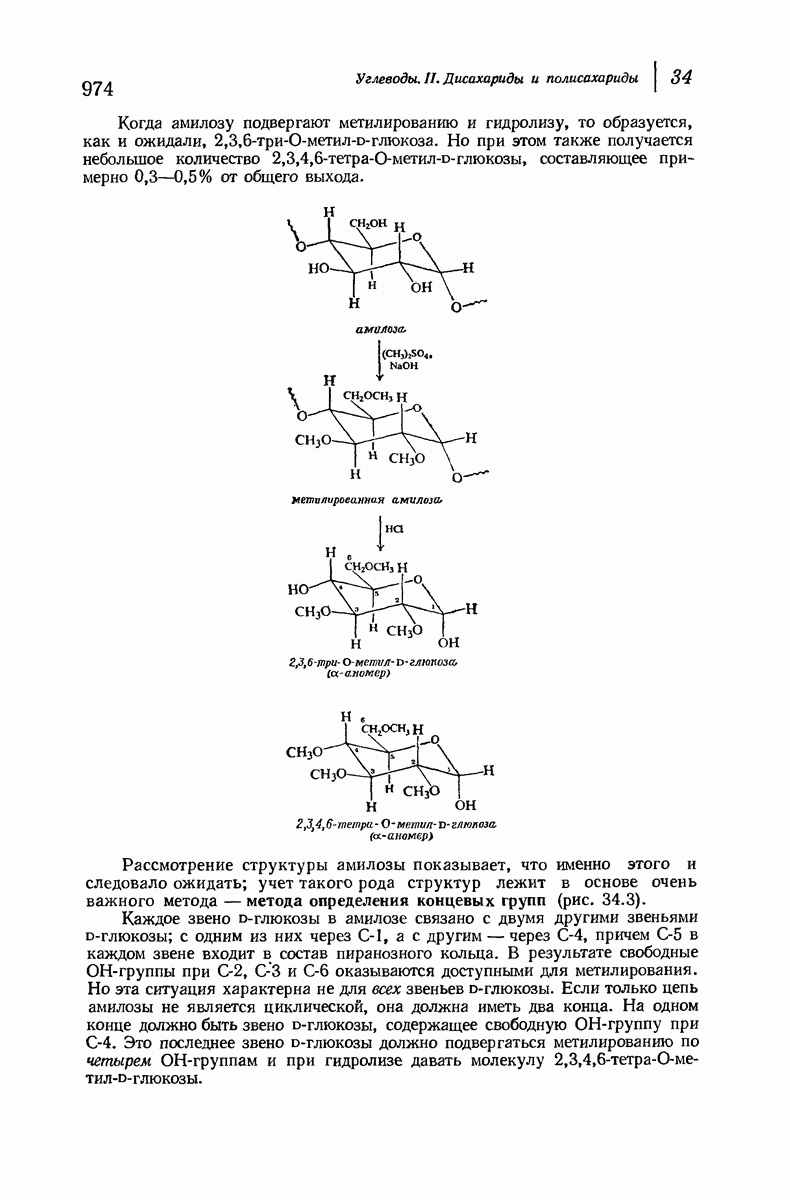
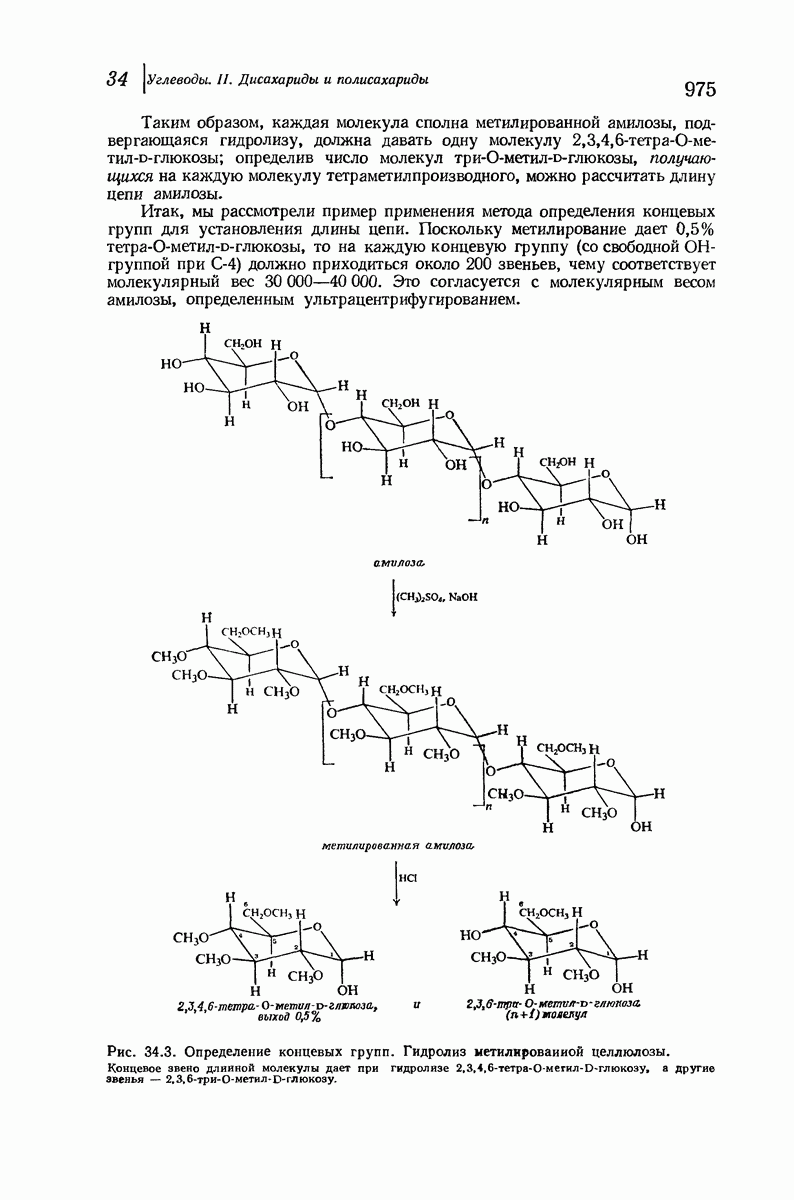
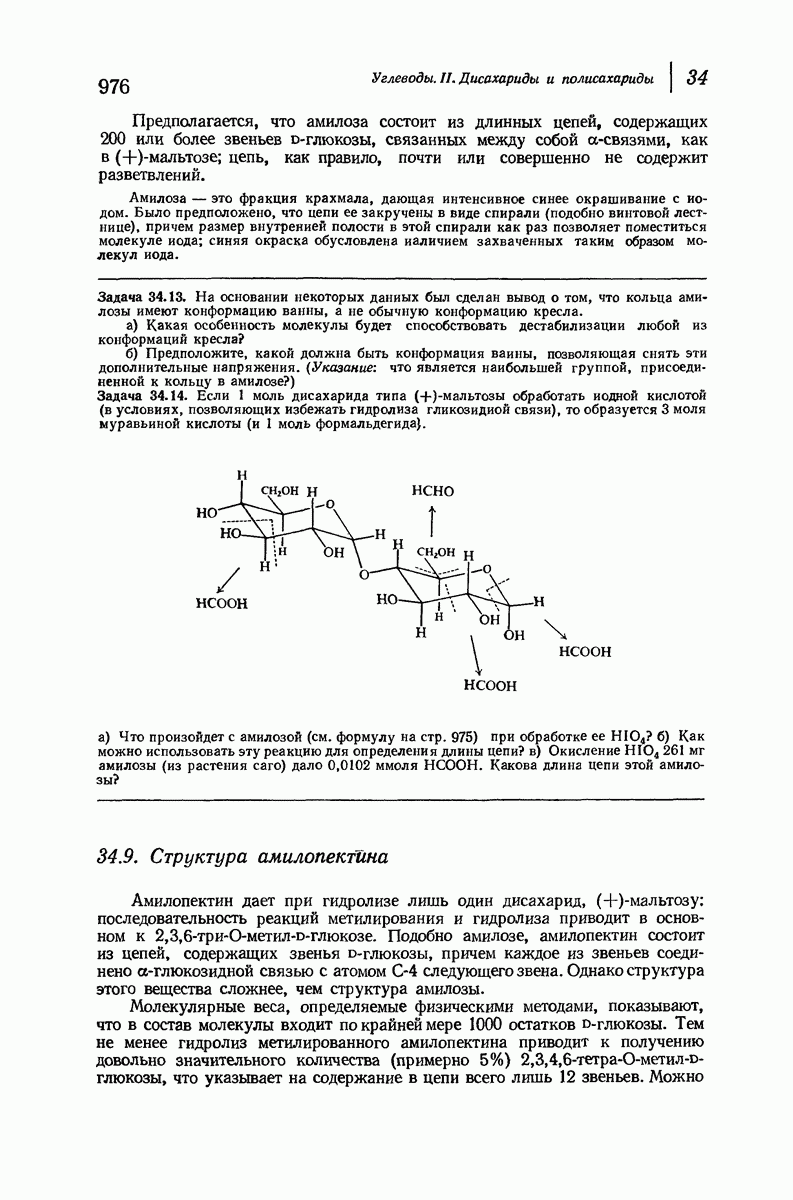
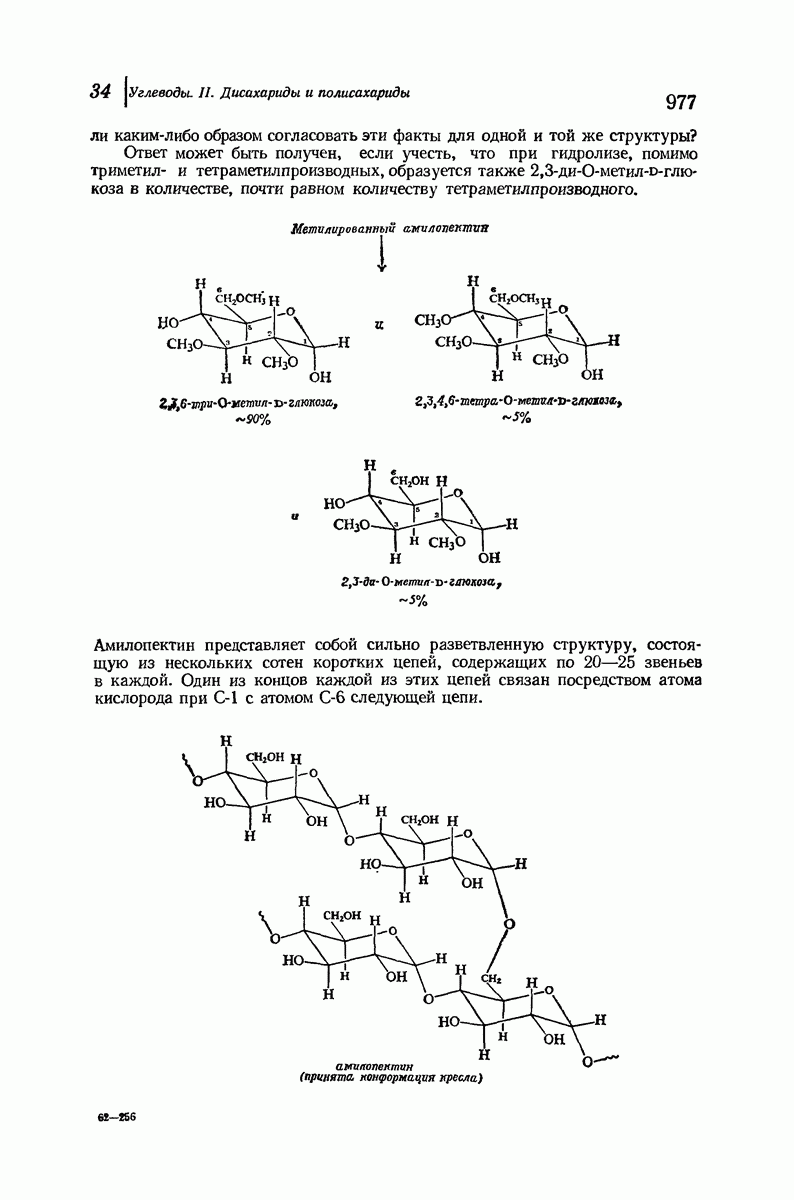
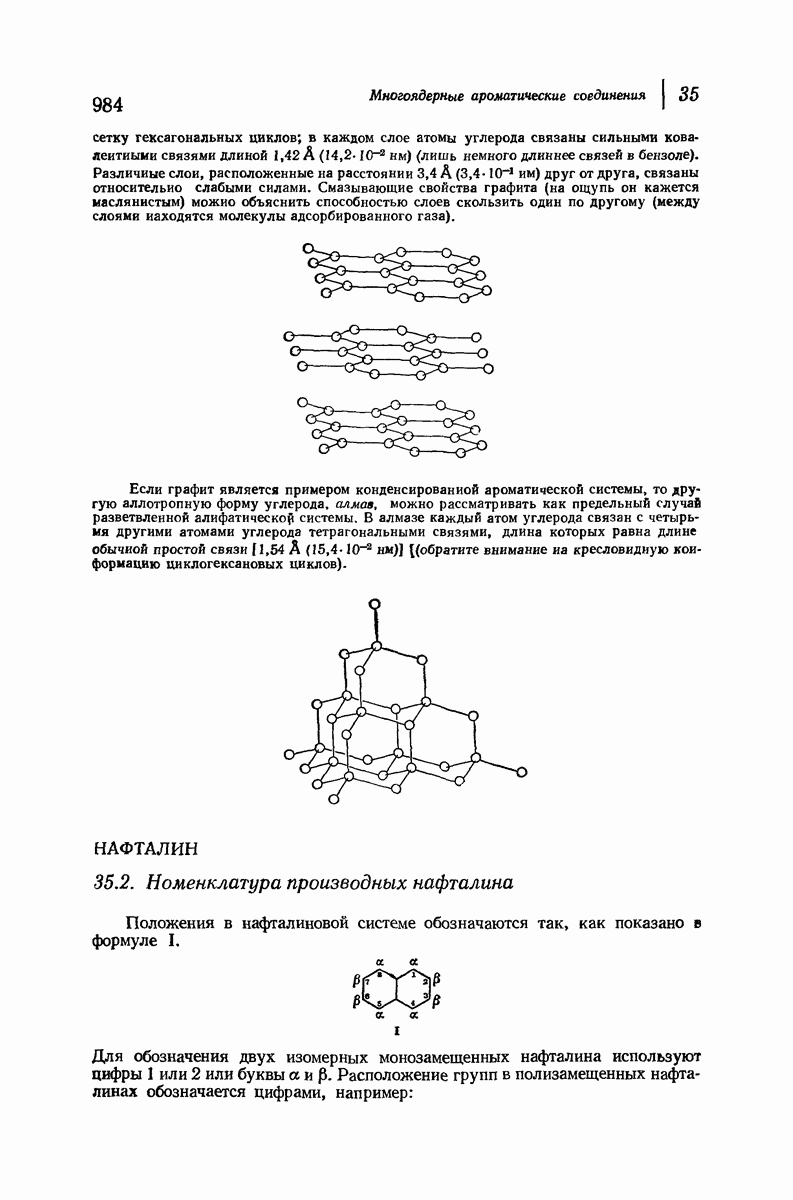
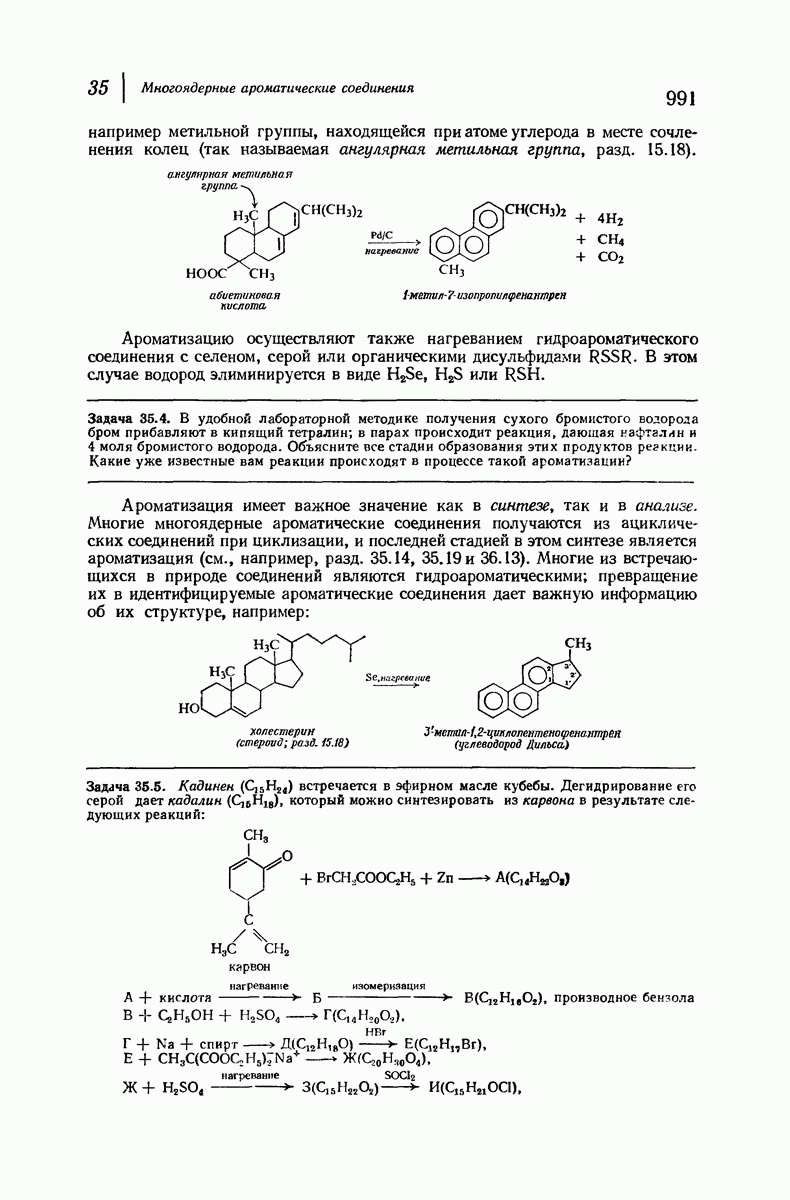
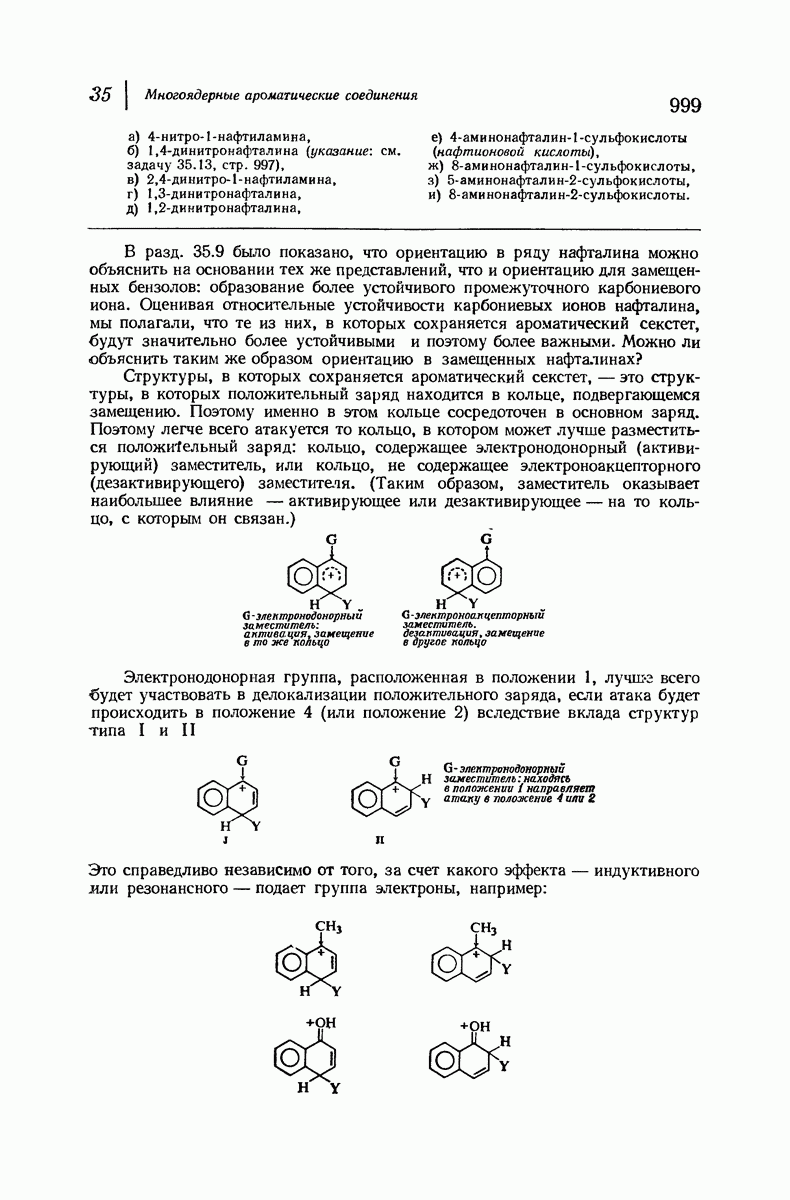
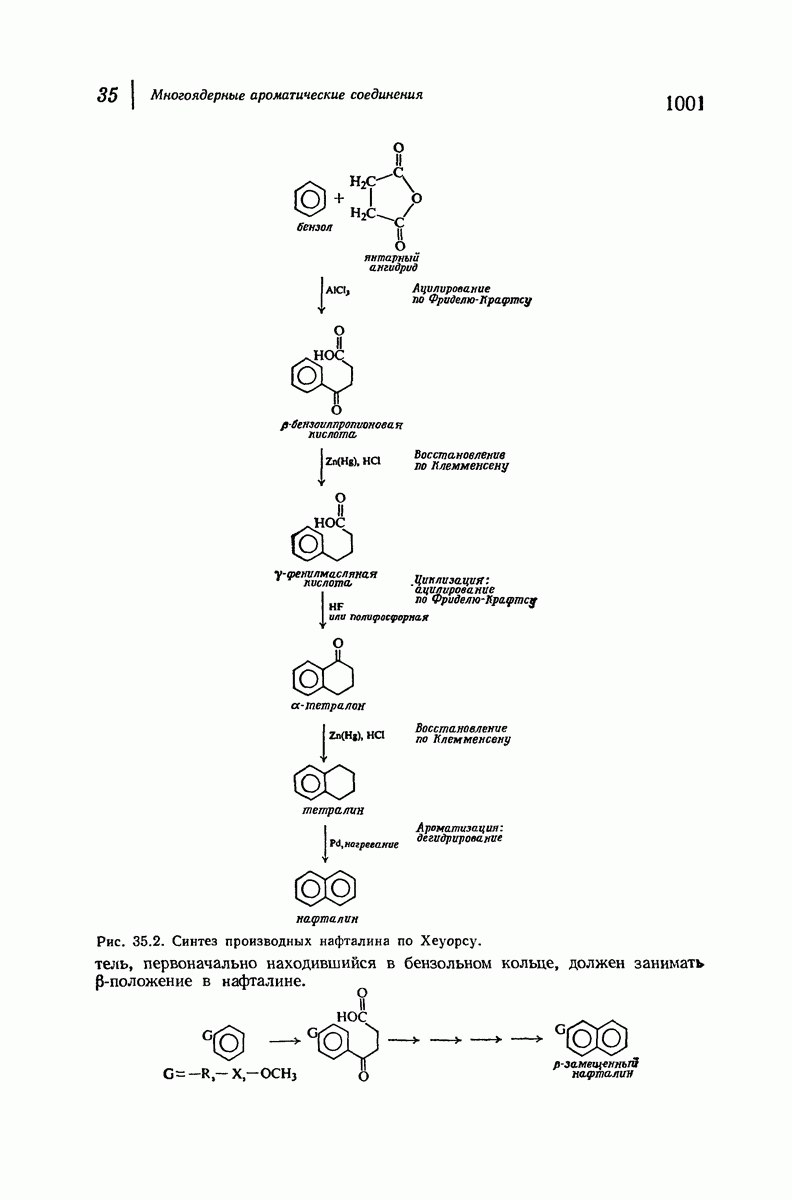
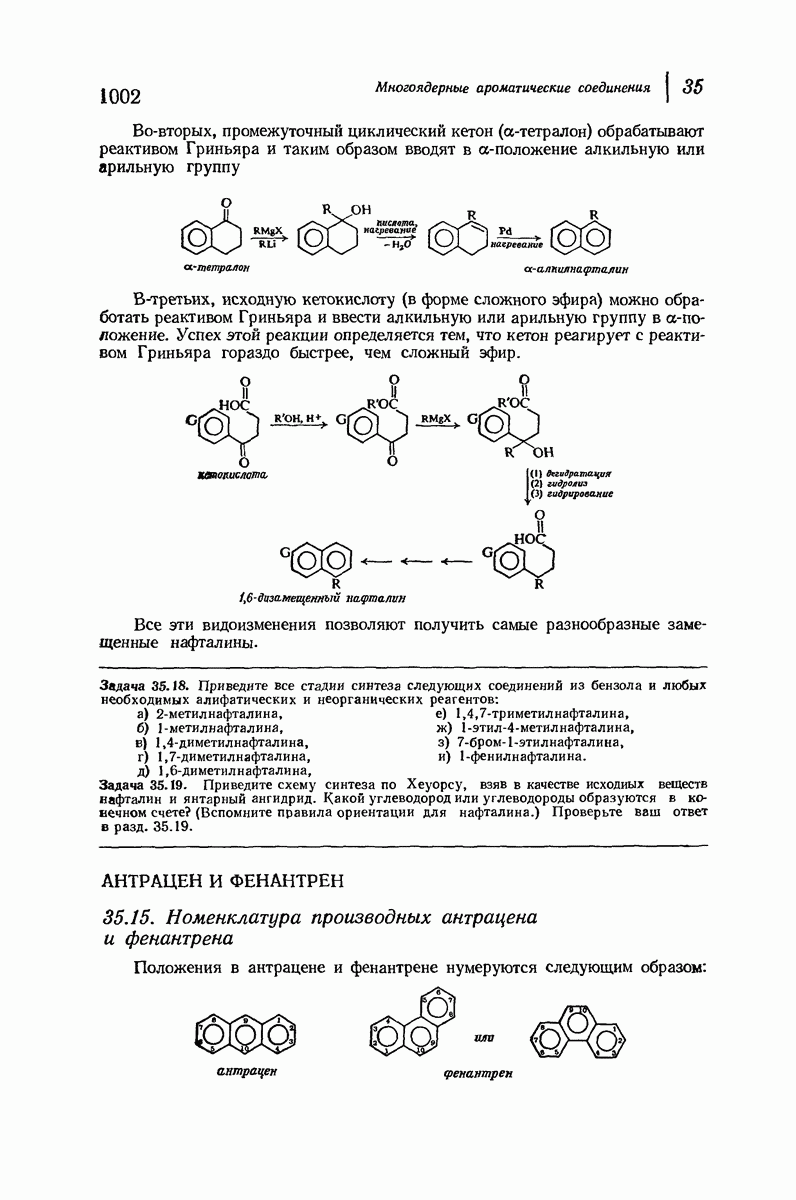
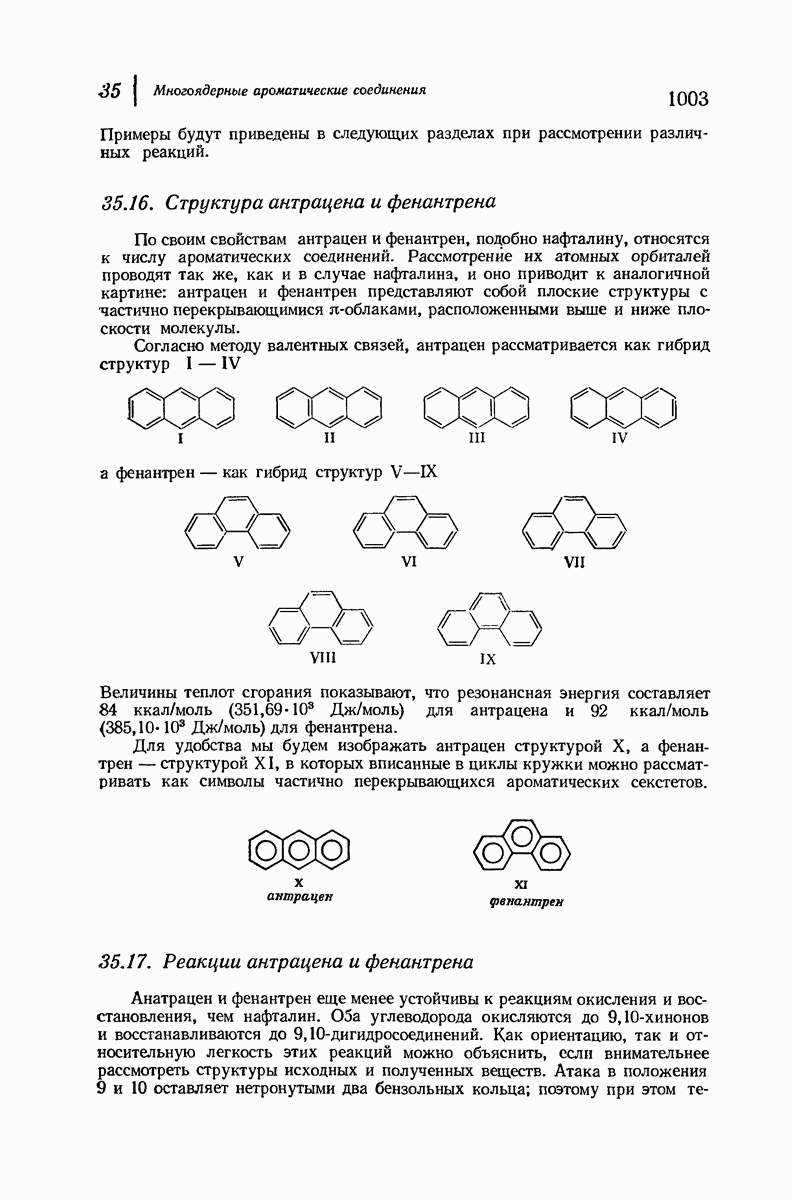
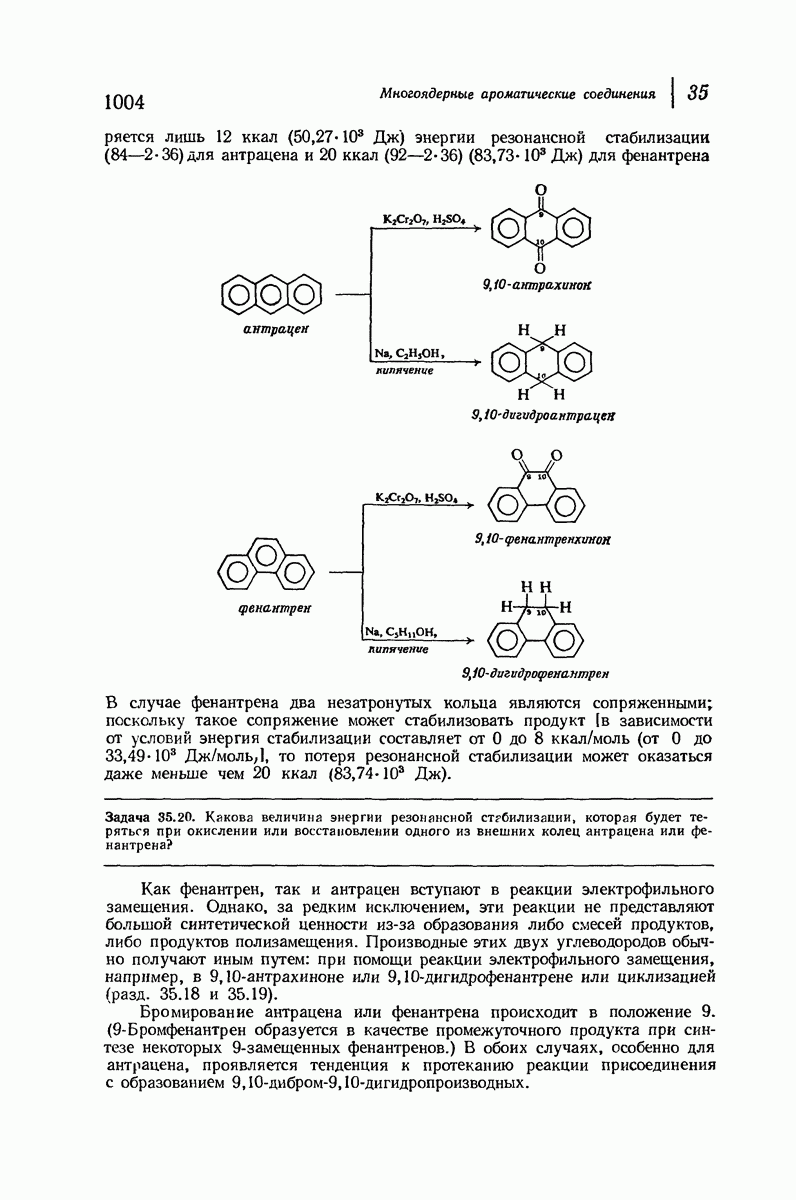
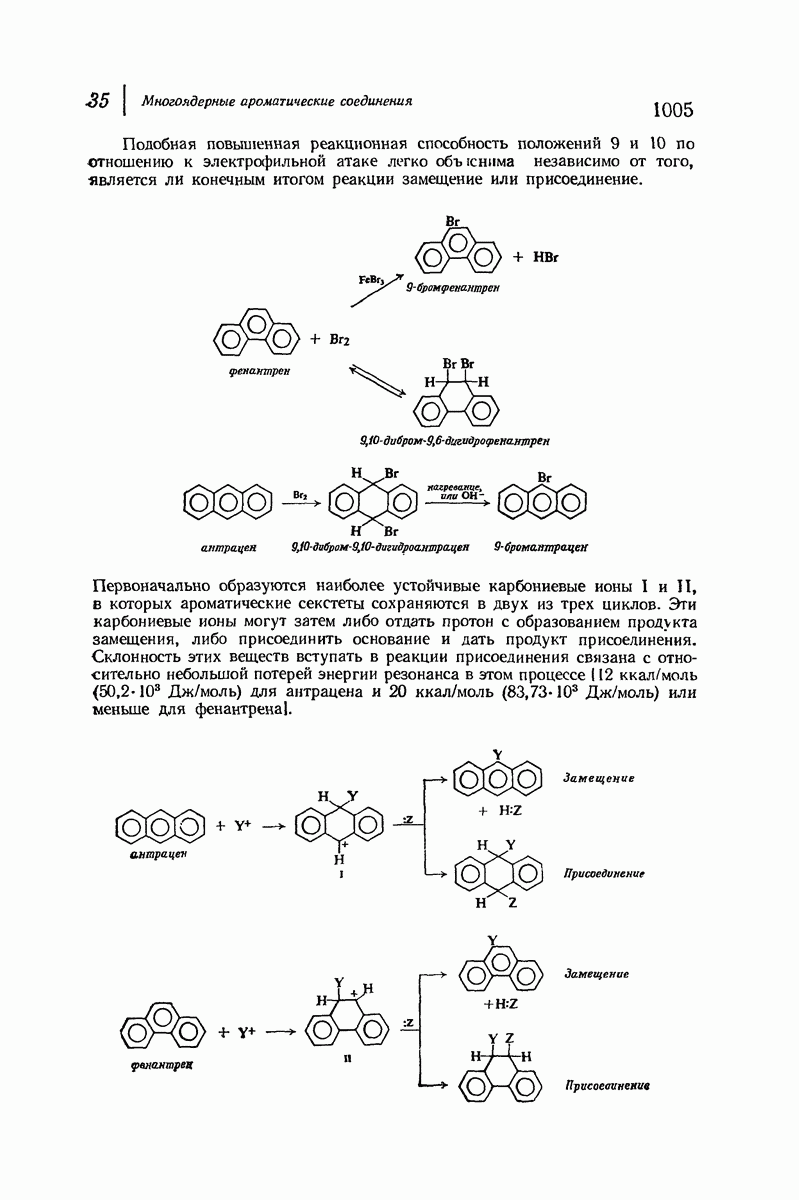
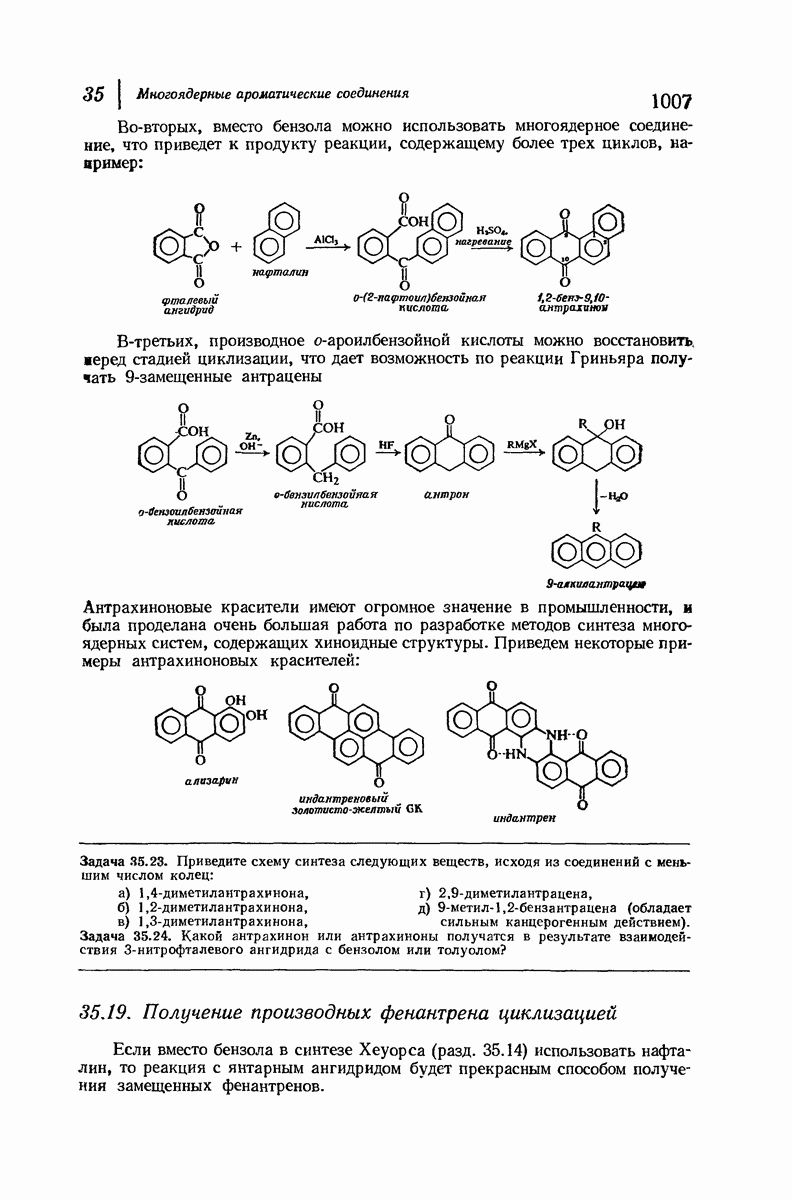
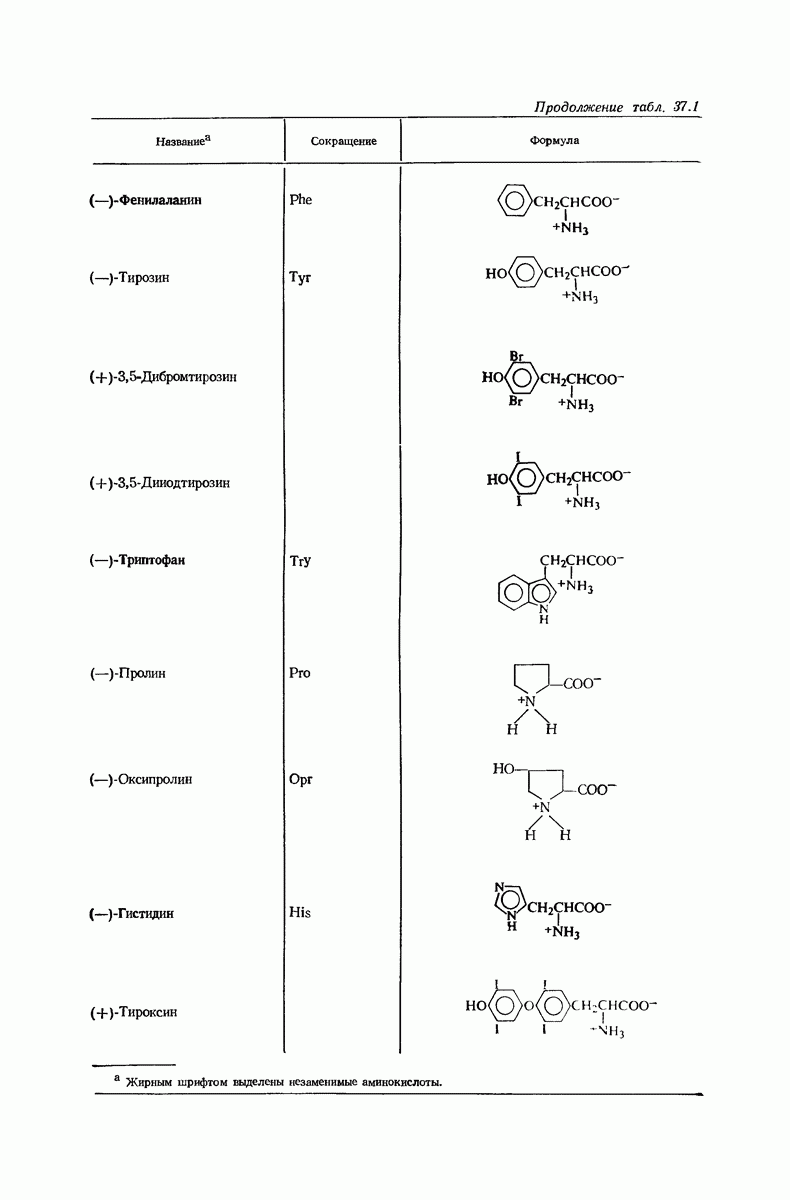
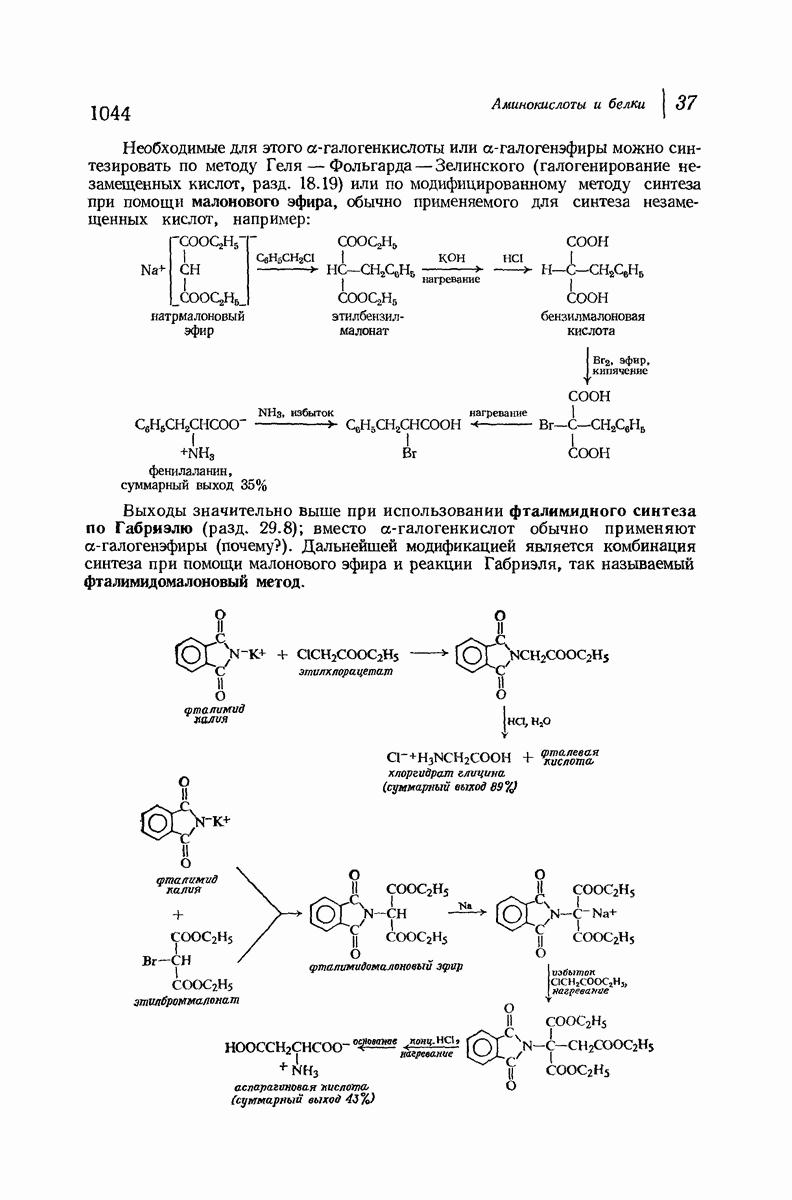
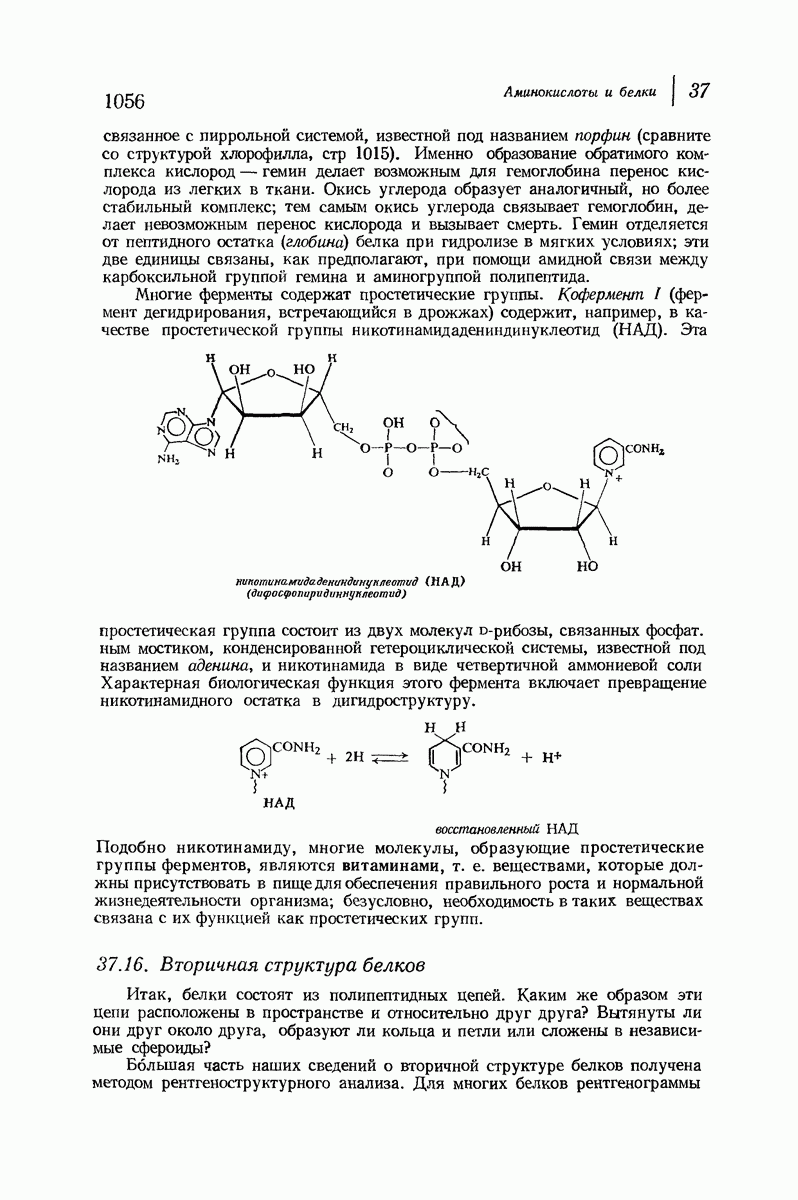
37.16. Вторичная структура белков
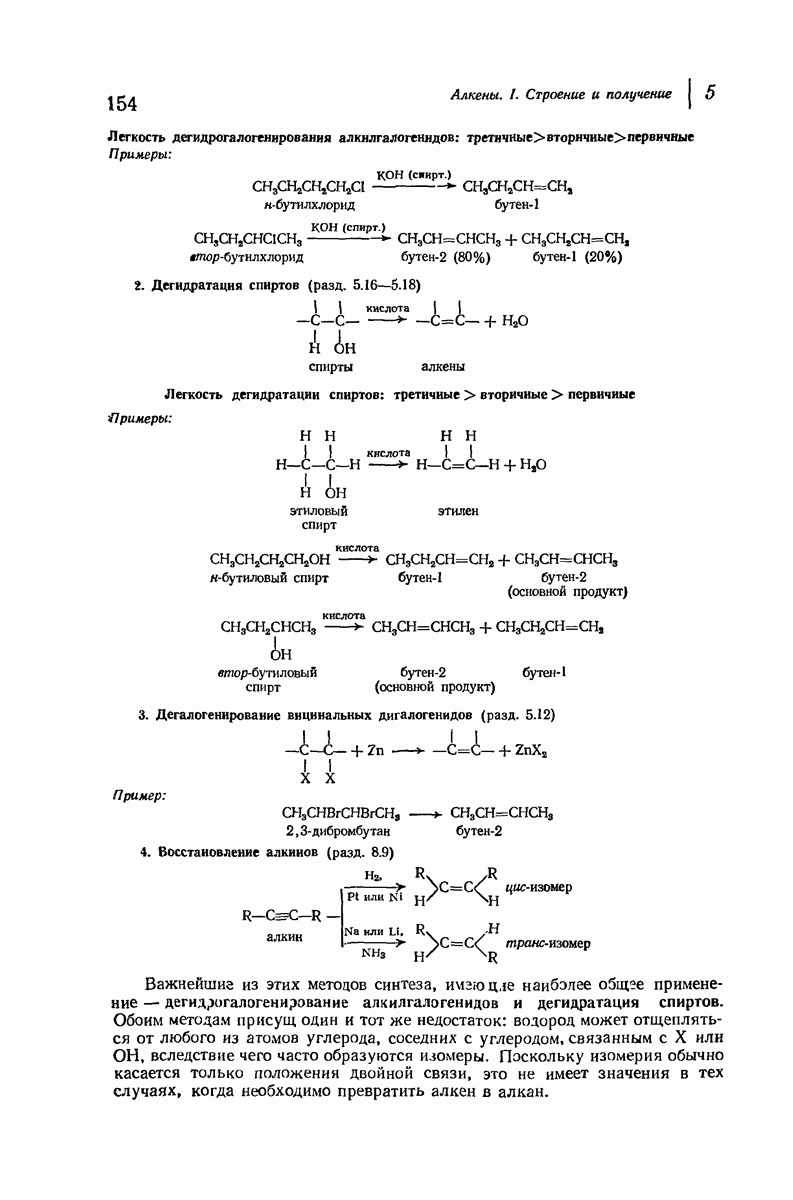
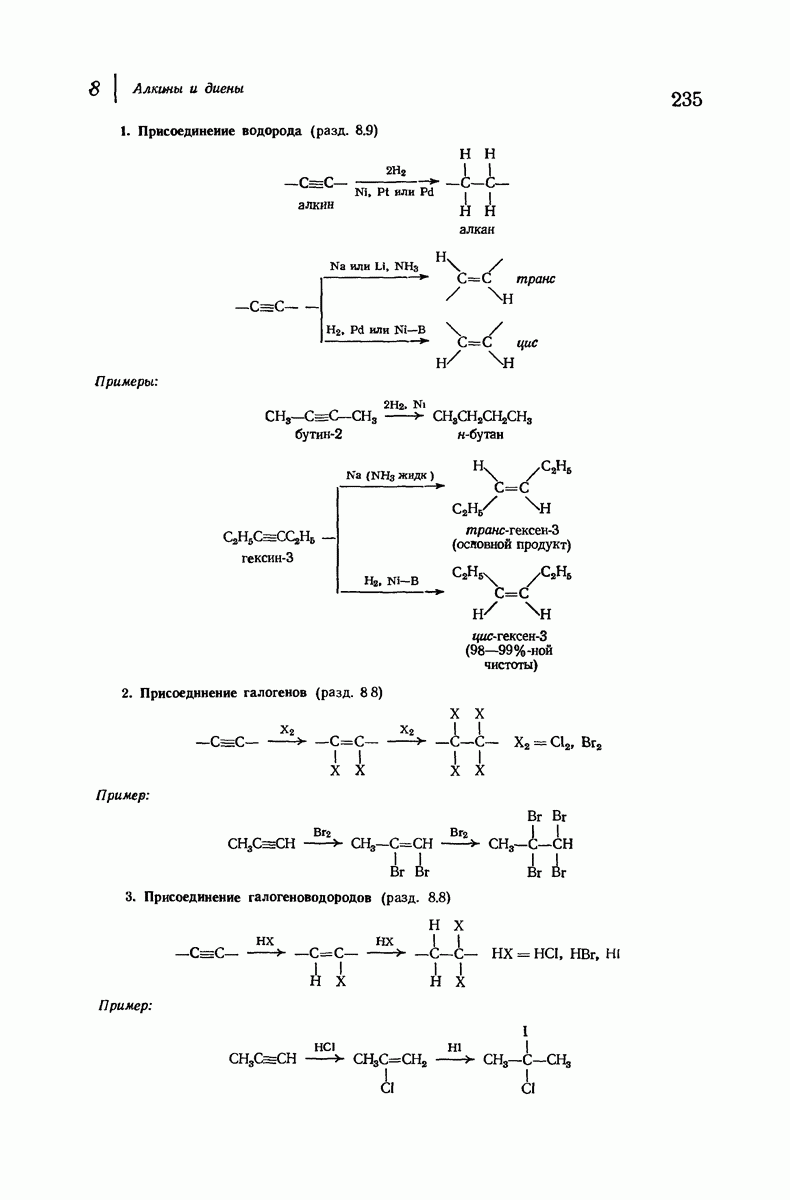
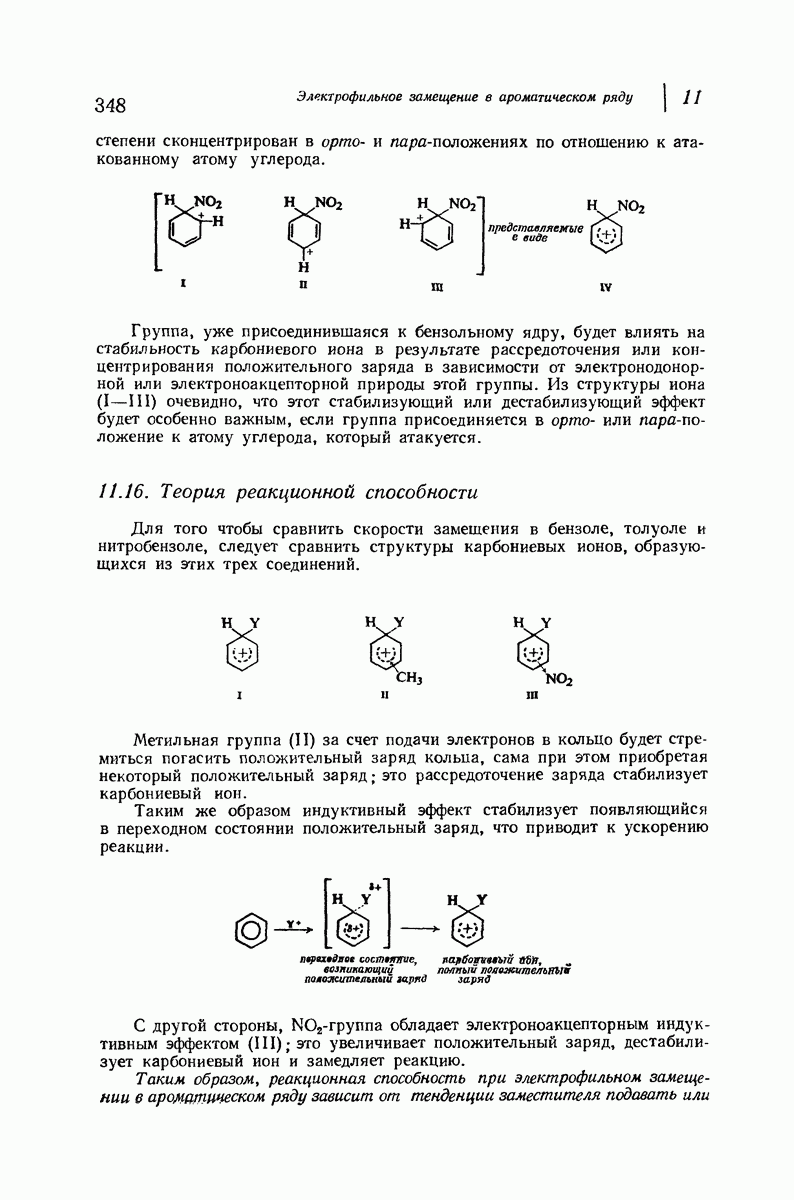
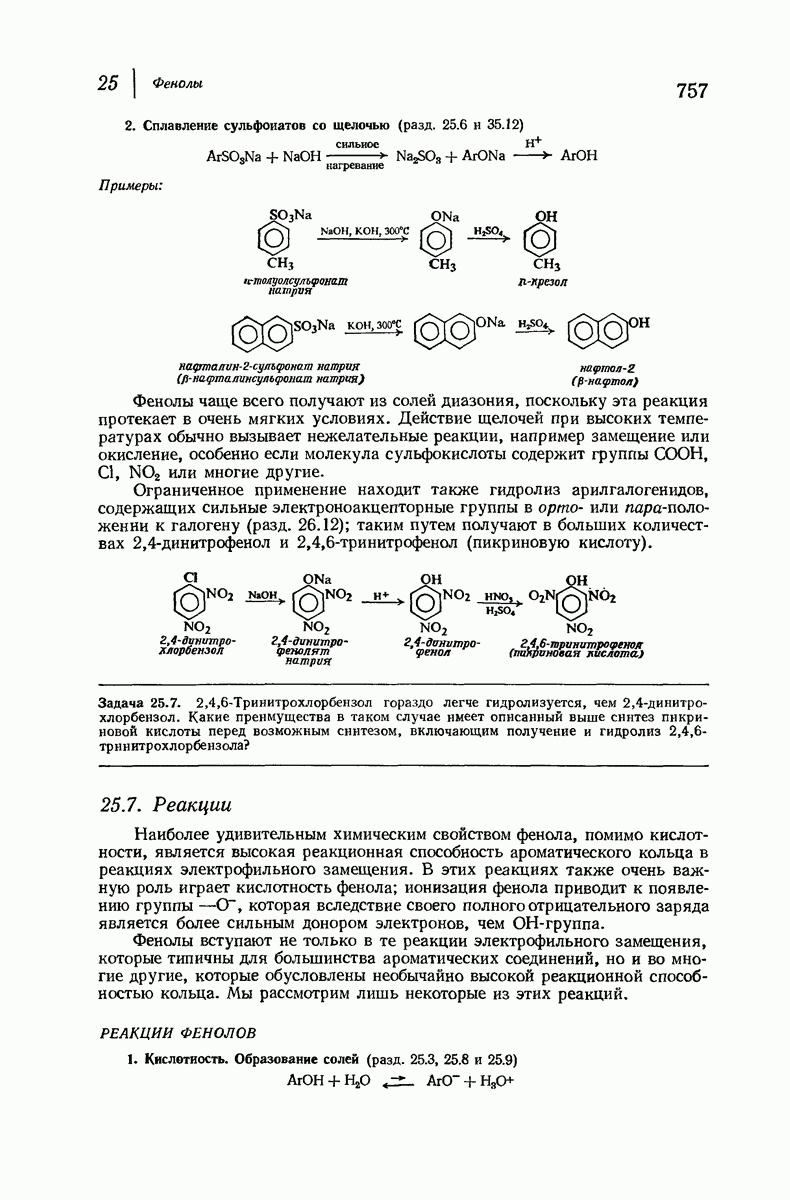
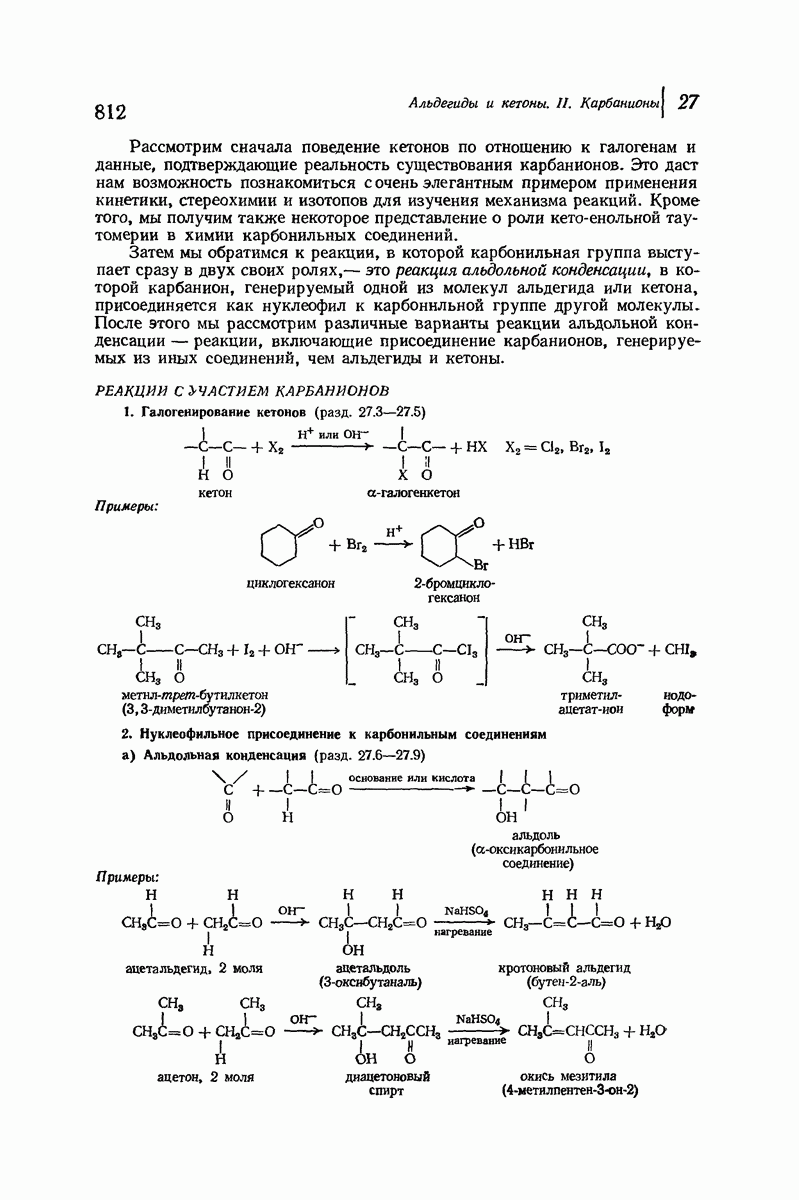
Итак, белки состоят из полипептидных цепей. Каким же образом эти цепи расположены в пространстве и относительно друг друга? Вытянуты ли они друг около друга, образуют ли кольца и петли или сложены в независимые сфероиды?
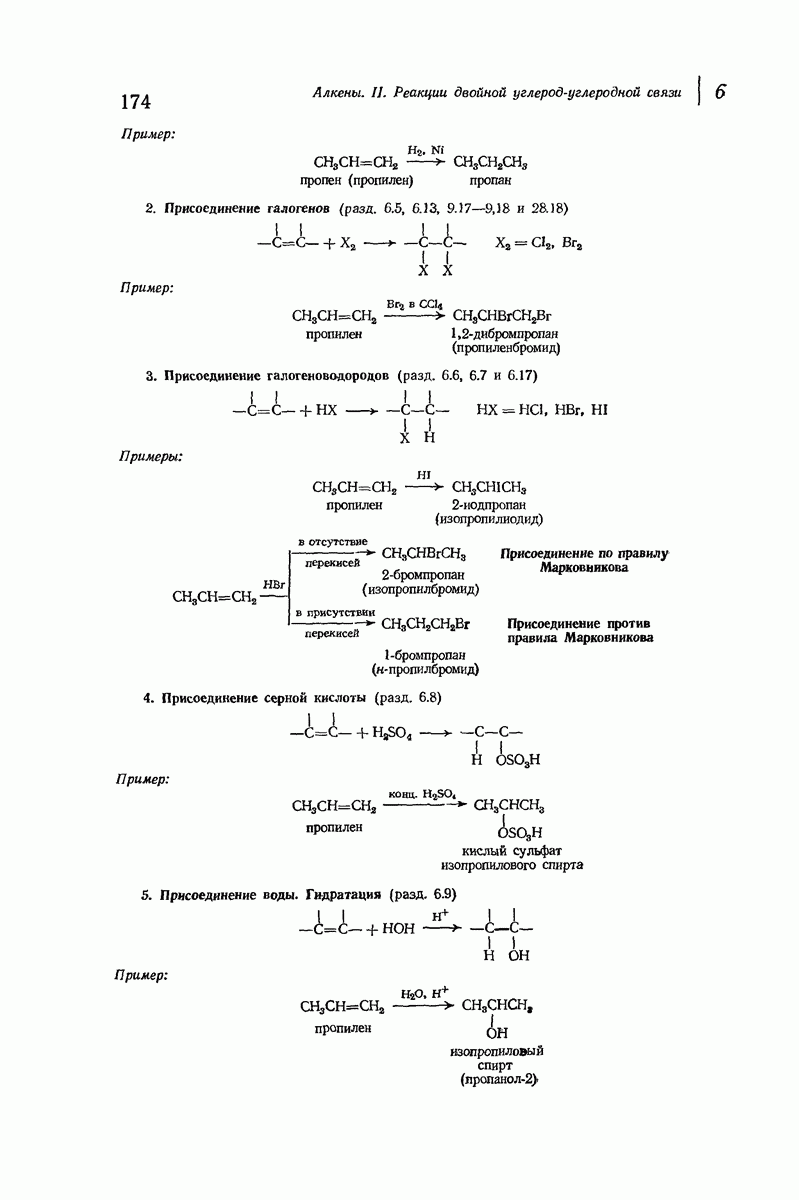
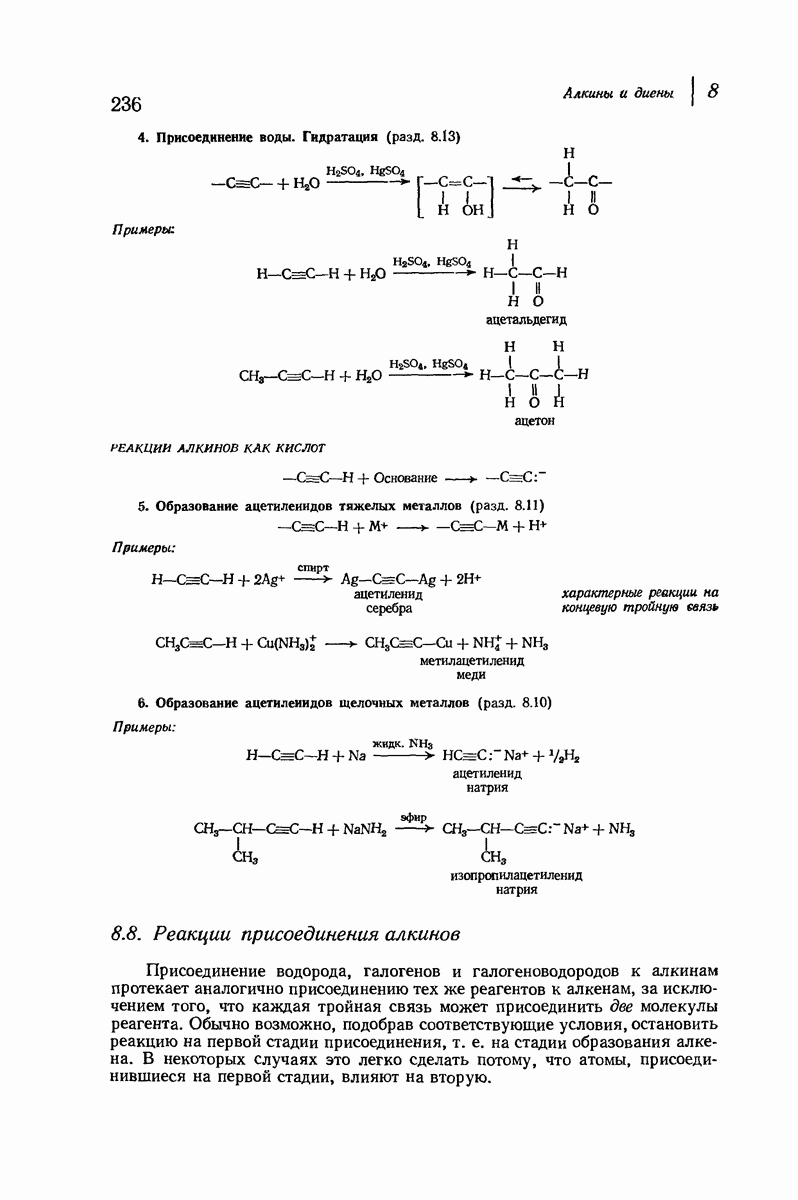
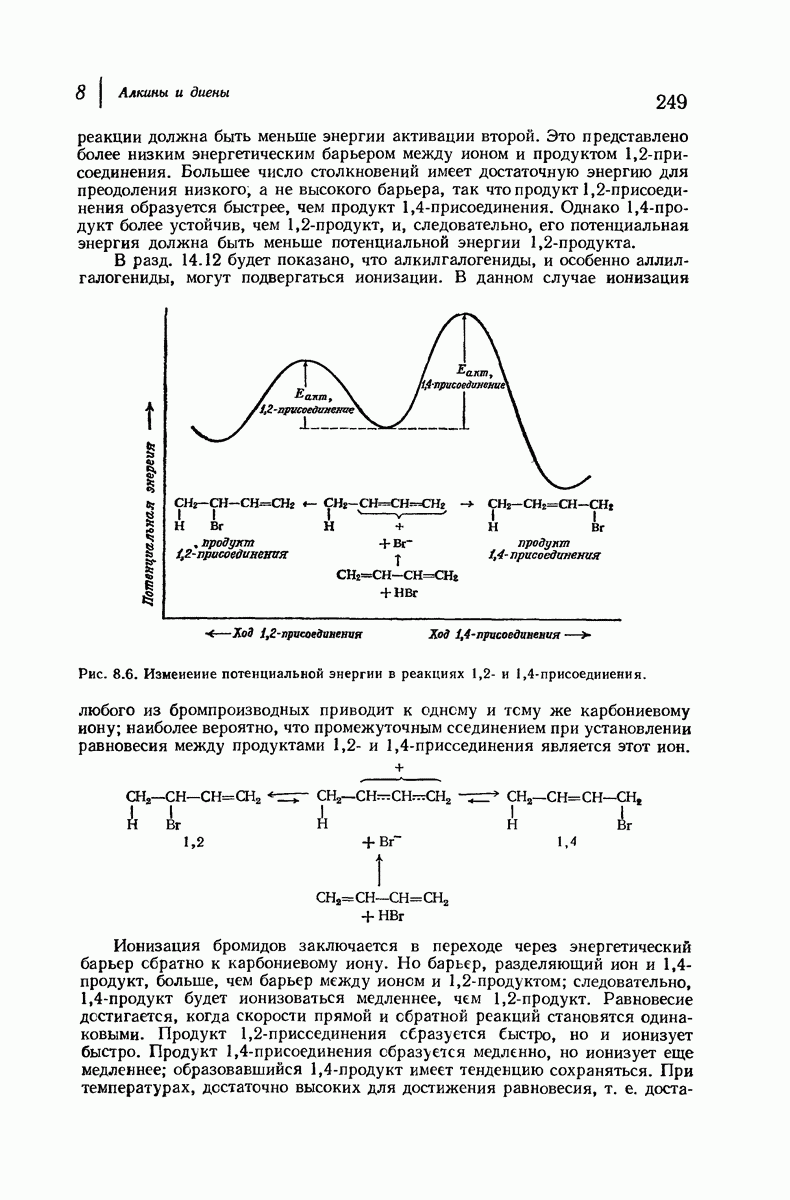
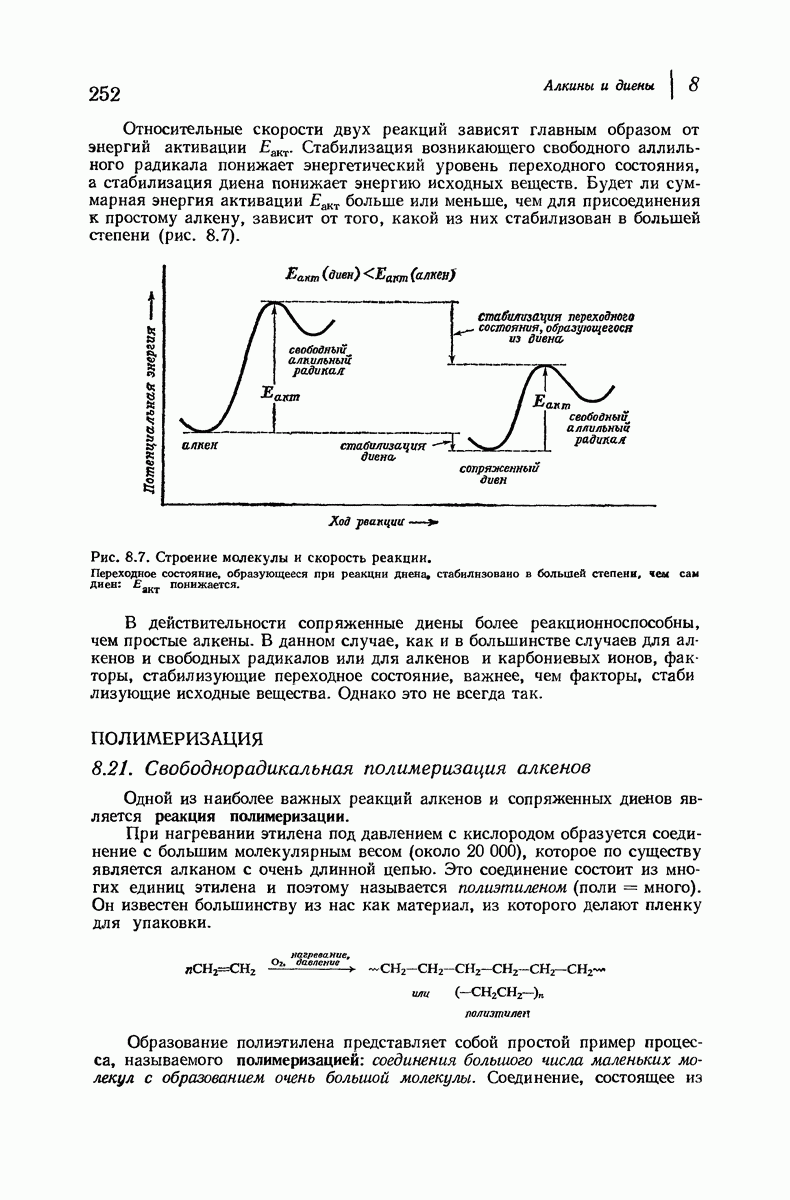
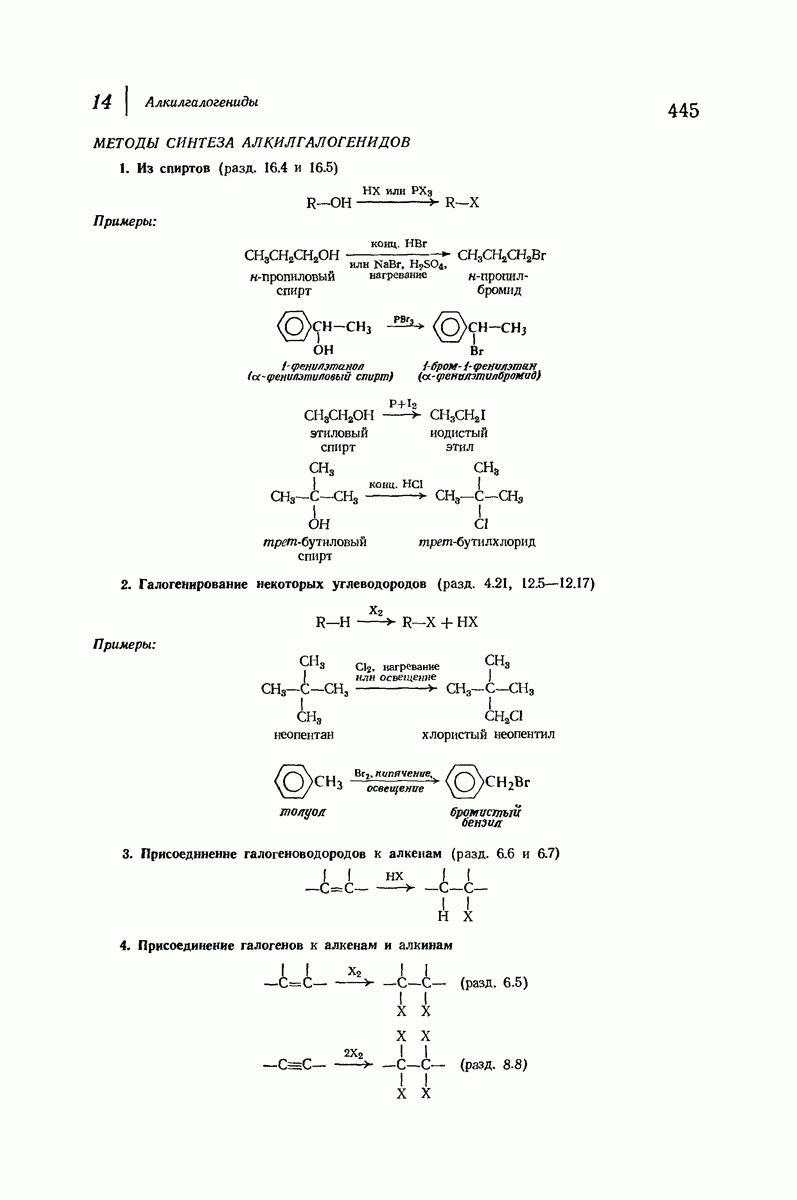
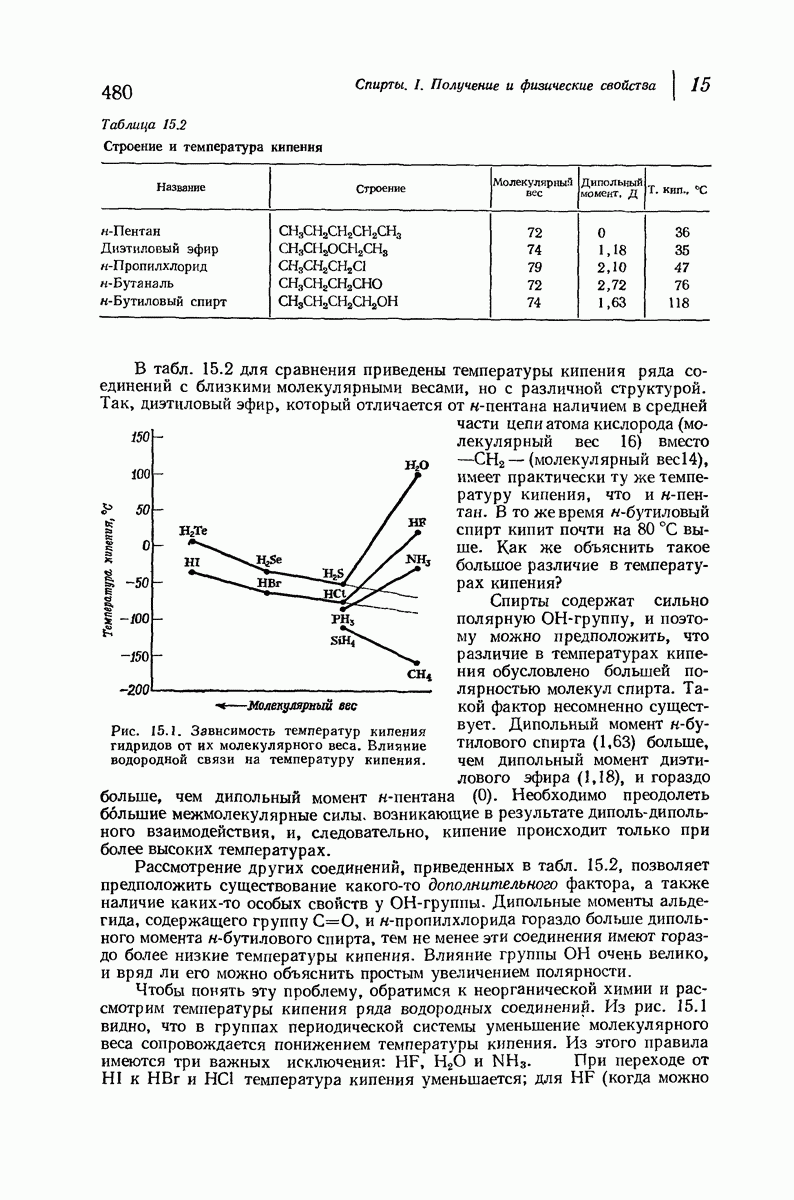
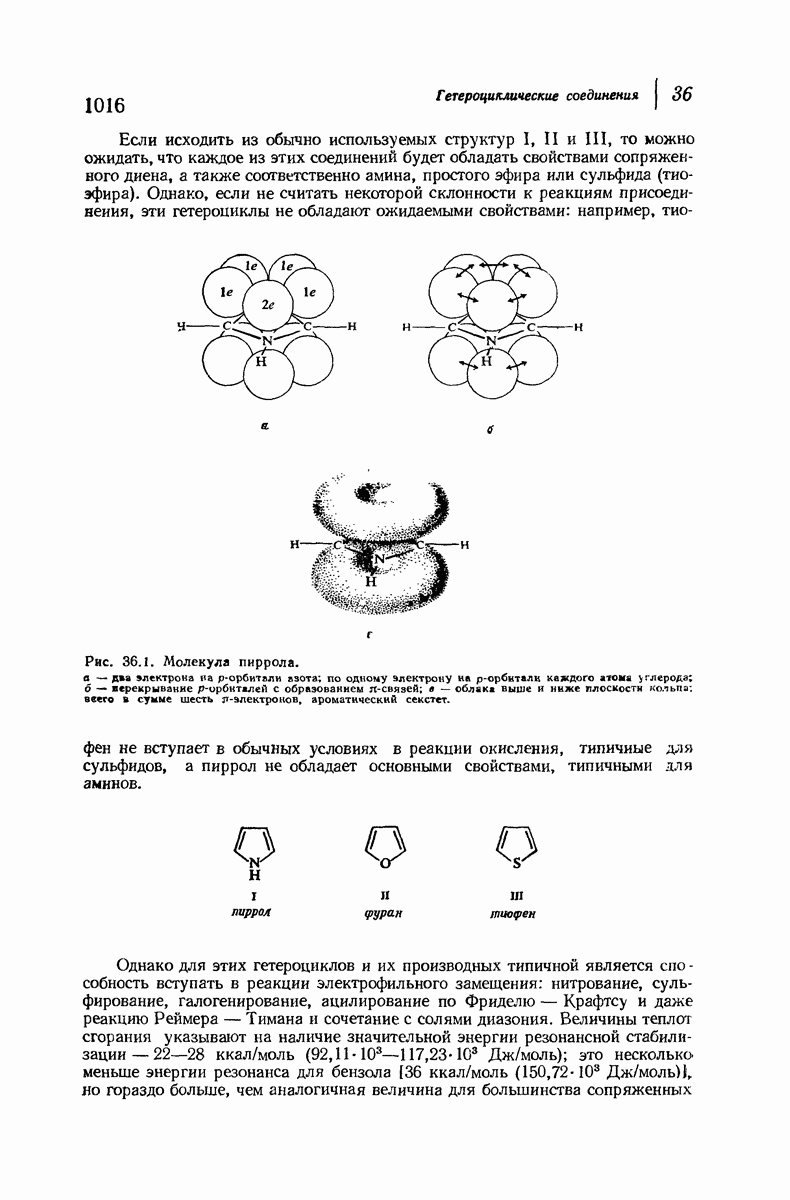
Большая часть наших сведений о вторичной структуре белков получена методом рентгеноструктурного анализа. Для многих белков рентгенограммы
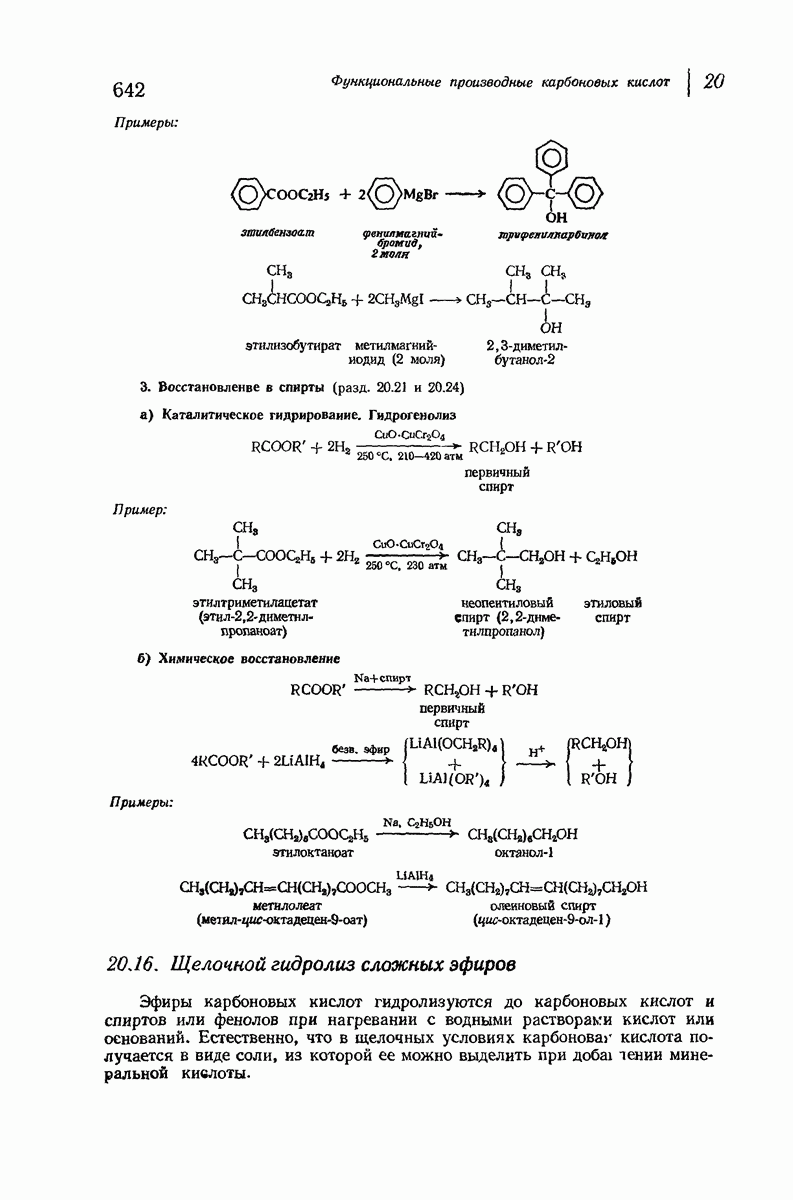
позволили обнаружить наличие правильных повторений некоторых структурных единиц. Например, для фиброина шелка расстояние между такими единицами (период идентичности) составляет 7,0 А  для
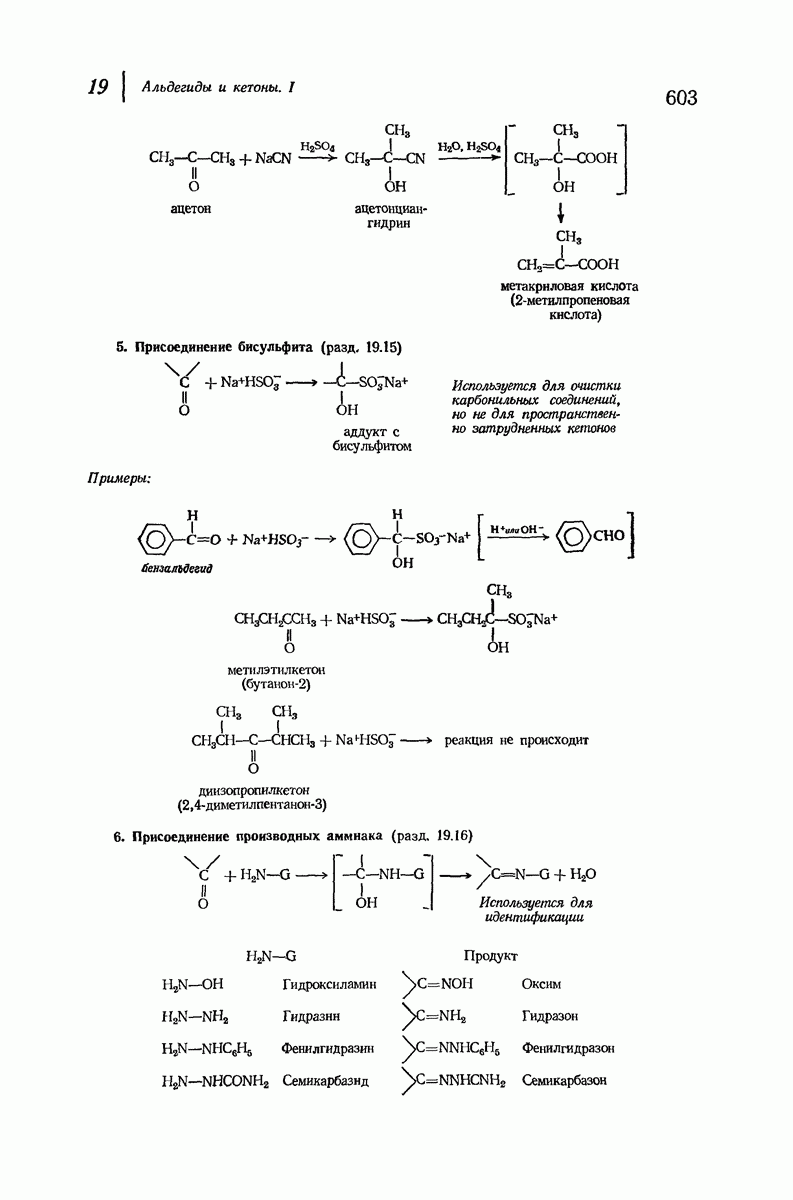
для  -кератина нерастянутой шерсти — 1,5 и 5,1 А
-кератина нерастянутой шерсти — 1,5 и 5,1 А 
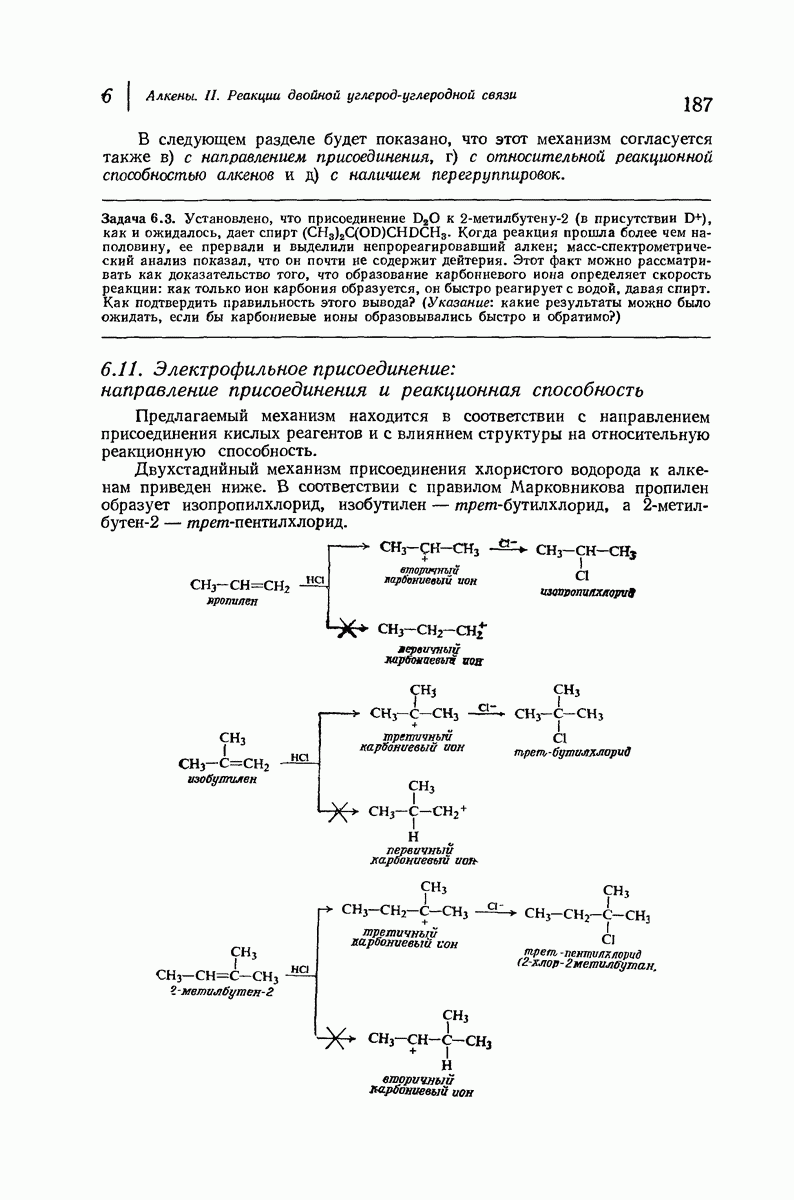
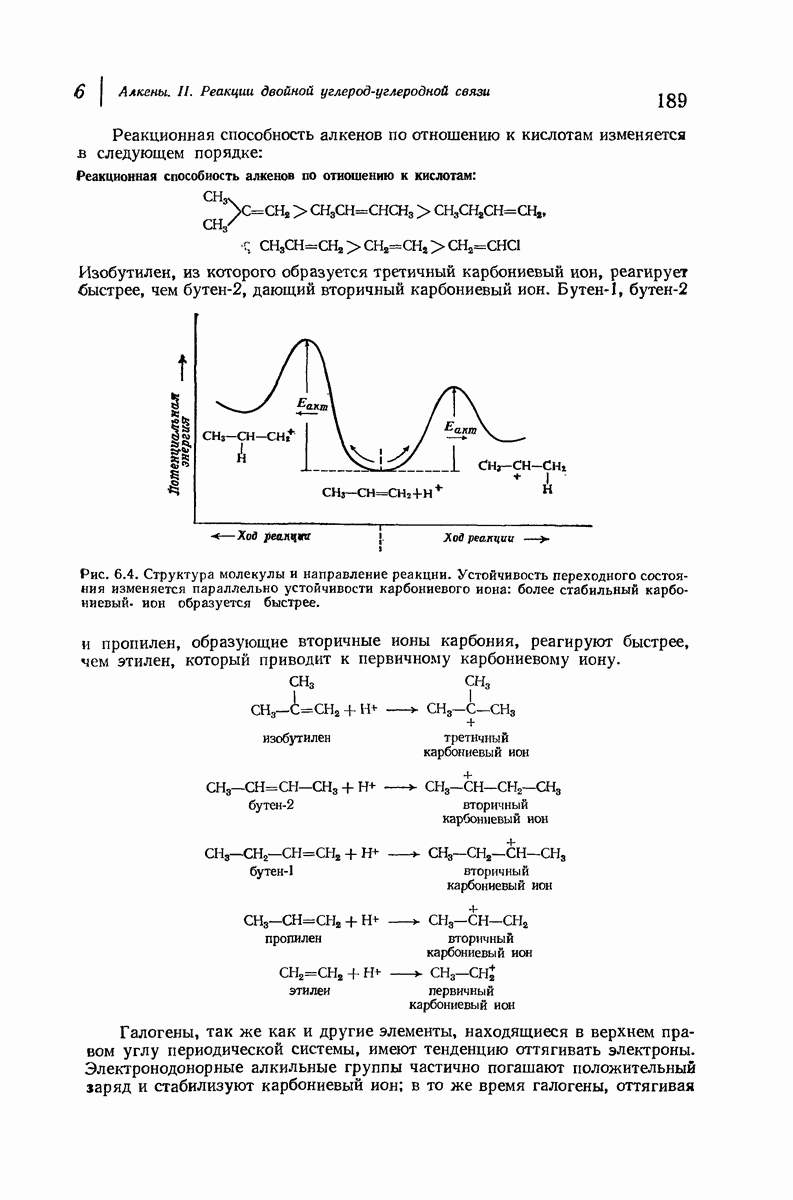
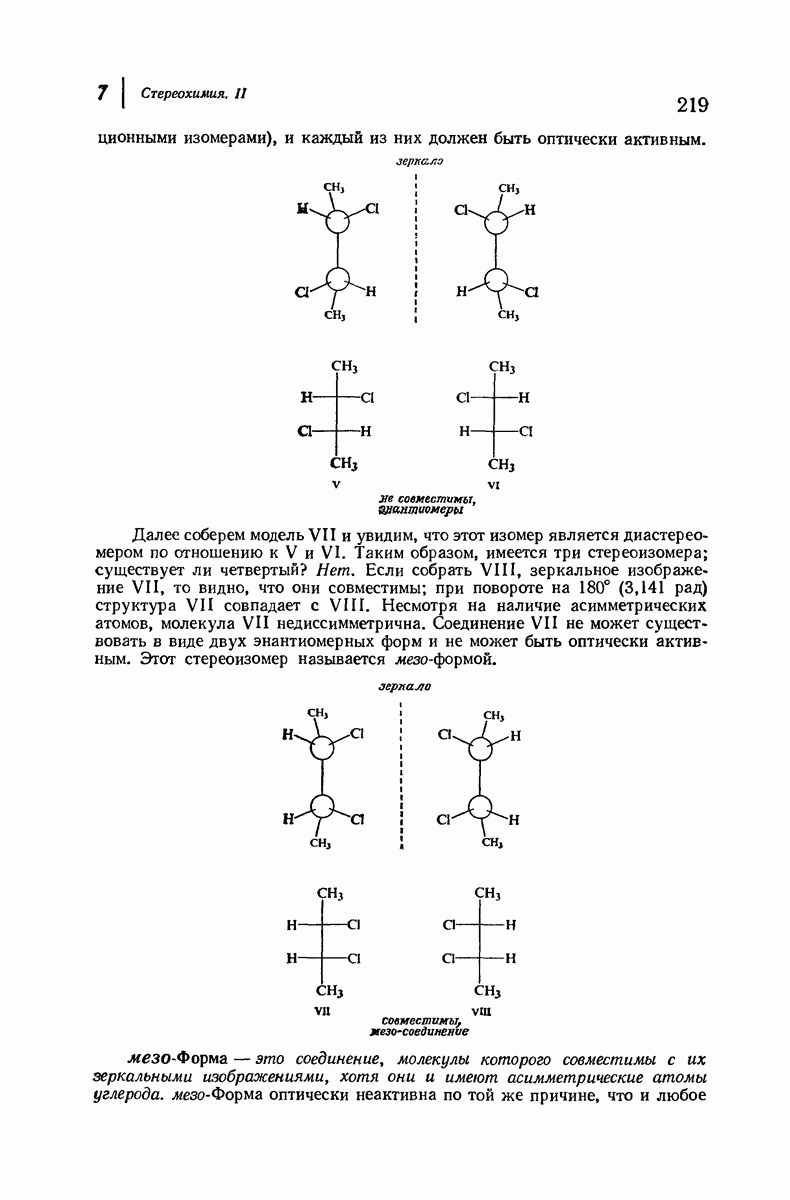
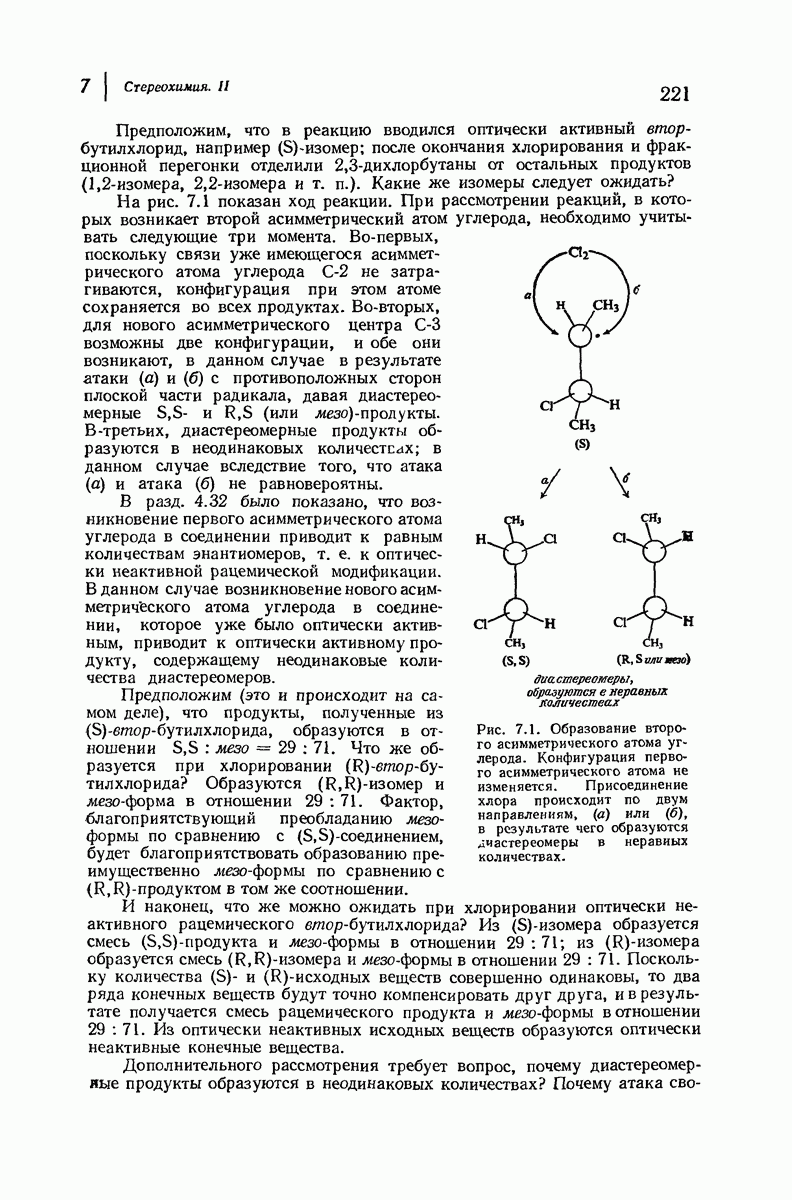
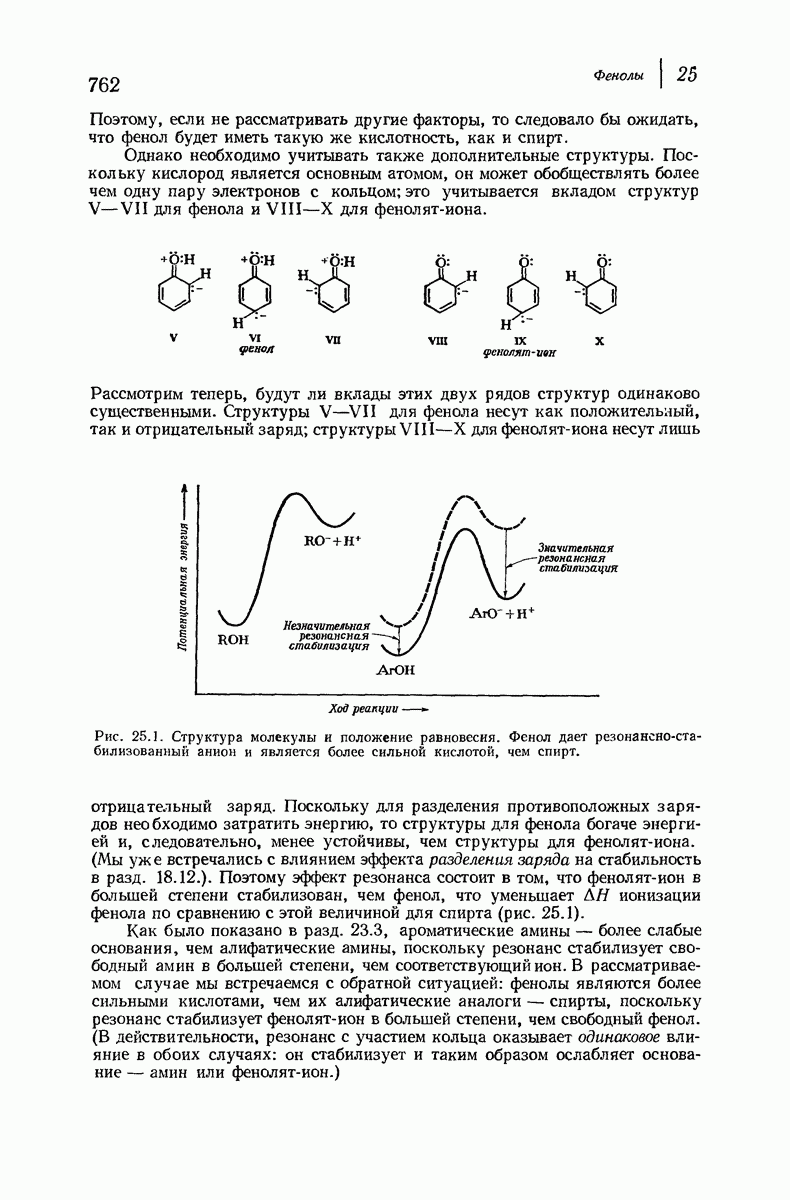
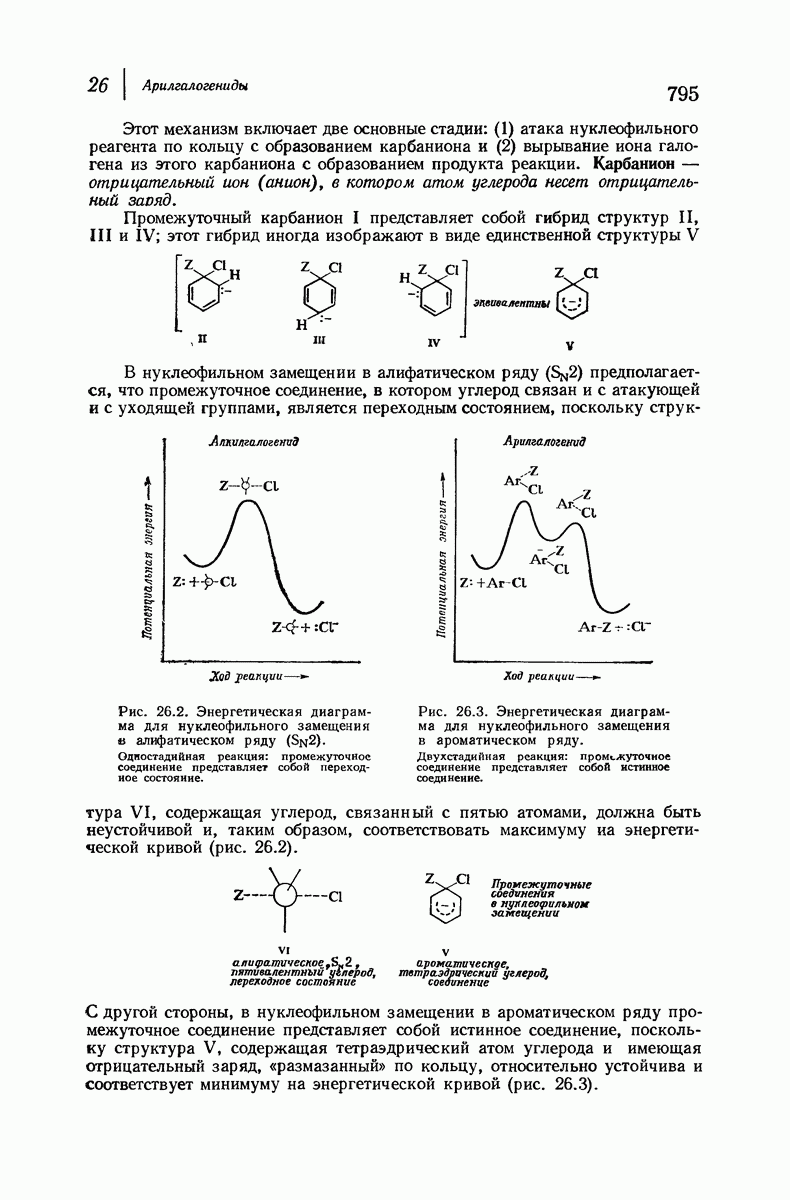
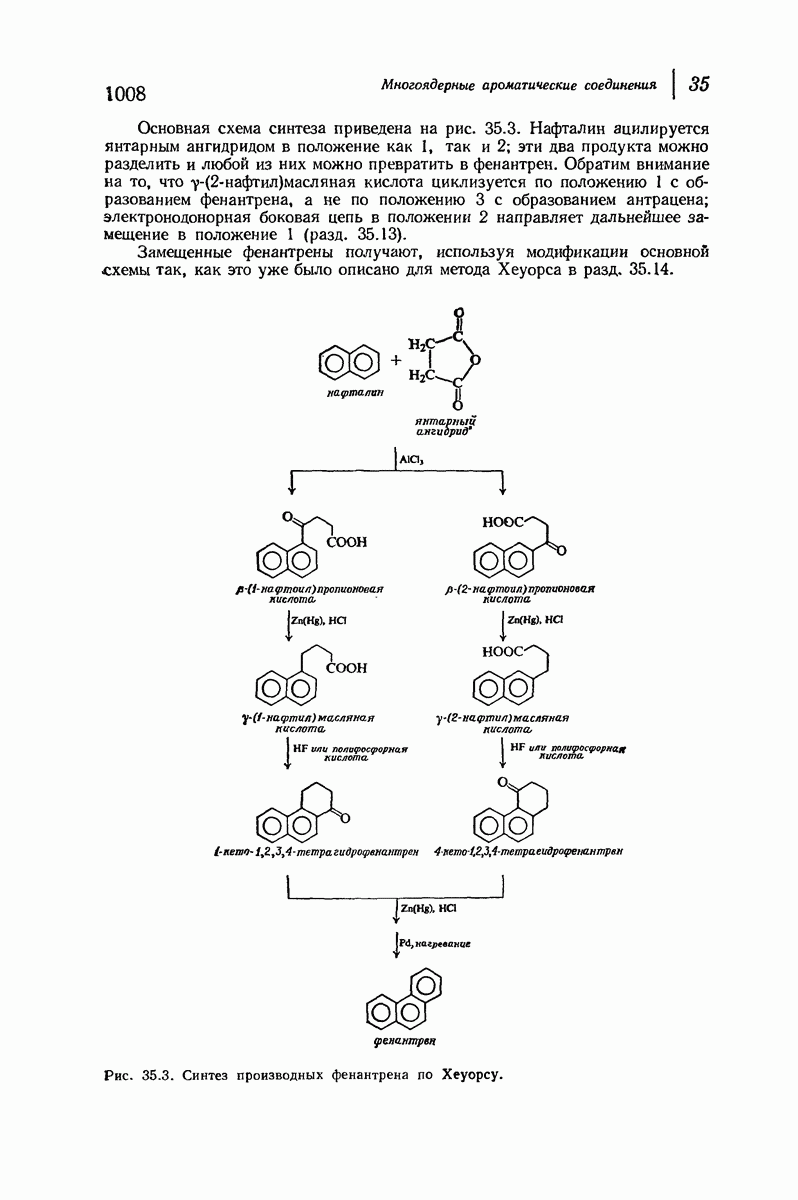
Дальнейшая задача состоит в построении структур, которые объясняют появление характеристичной картины дифракции рентгеновских лучей и в то же время согласуются с такими сведениями о первичной структуре, как данные о длинах и углах связей, плоское строение амидной группы, сходство конфигураций всех звеньев относительно асимметрического атома углерода (все  -ряда), размер и последовательность расположения боковых групп. Ключевым моментом при решении этой задачи является учет стабилизующего влияния водородных связей (5—10 ккал/моль на одну водородную связь) и того принципа, что наиболее устойчивой структурой будет структура, допускающая образование максимального числа водородных связей. Кроме того, на основании данных по изучению более простых соединений принимается, что связь
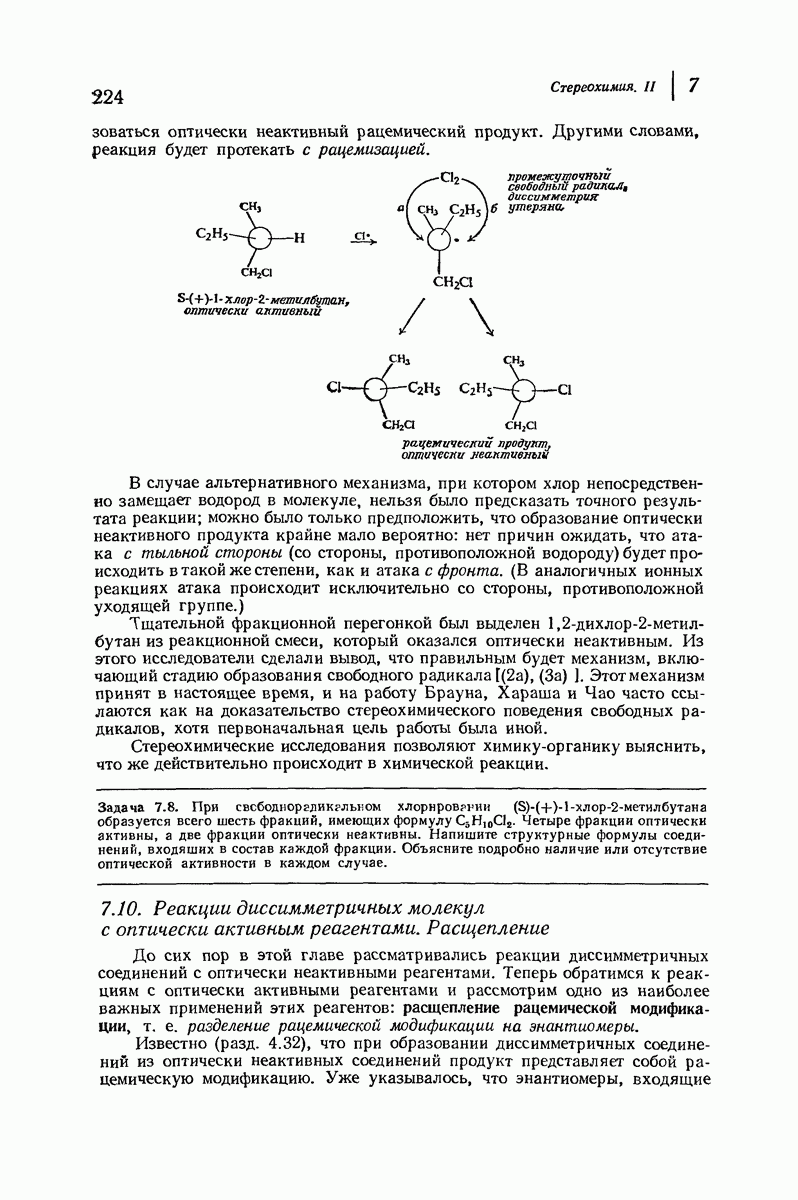
-ряда), размер и последовательность расположения боковых групп. Ключевым моментом при решении этой задачи является учет стабилизующего влияния водородных связей (5—10 ккал/моль на одну водородную связь) и того принципа, что наиболее устойчивой структурой будет структура, допускающая образование максимального числа водородных связей. Кроме того, на основании данных по изучению более простых соединений принимается, что связь  почти линейна и водород расположен на линии связи между кислородом и азотом или отклоняется от нее не более чем на 20°. Во всей этой работе очень полезным оказалось изучение более простых синтетических полипептидов, содержащих аминокислотные остатки одного вида.
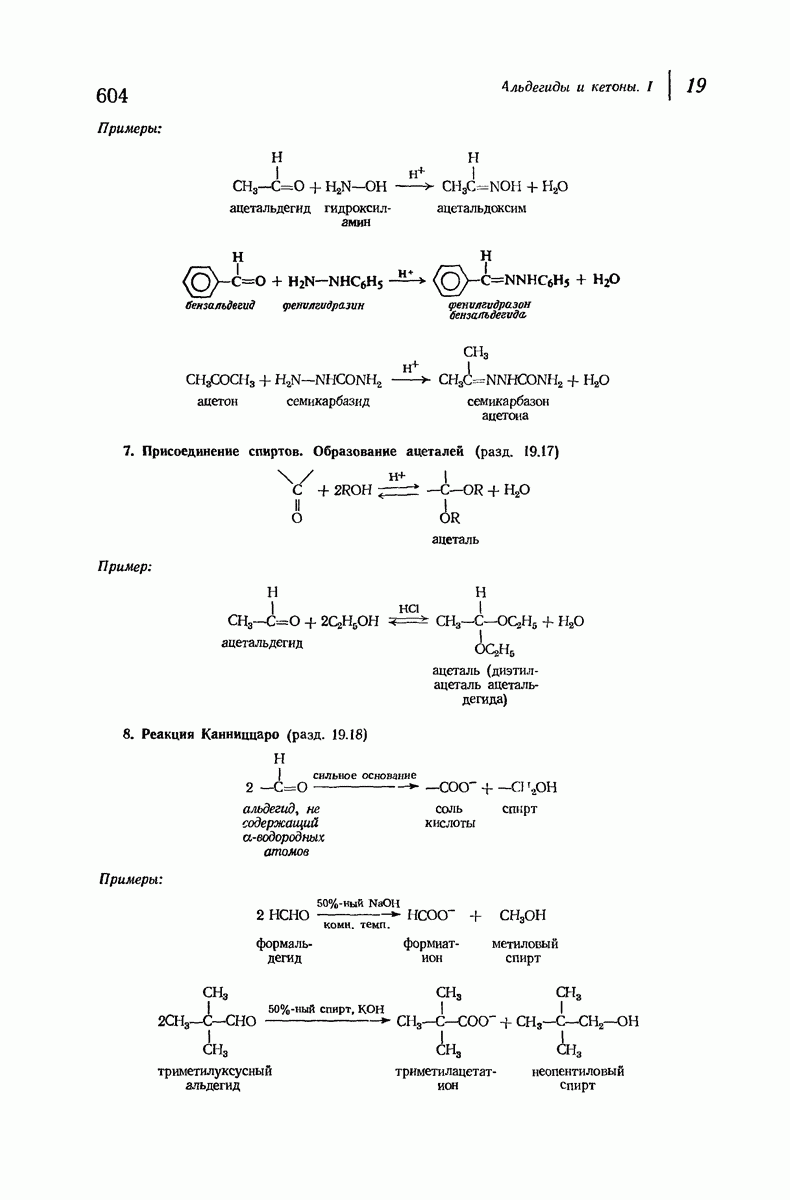
почти линейна и водород расположен на линии связи между кислородом и азотом или отклоняется от нее не более чем на 20°. Во всей этой работе очень полезным оказалось изучение более простых синтетических полипептидов, содержащих аминокислотные остатки одного вида.
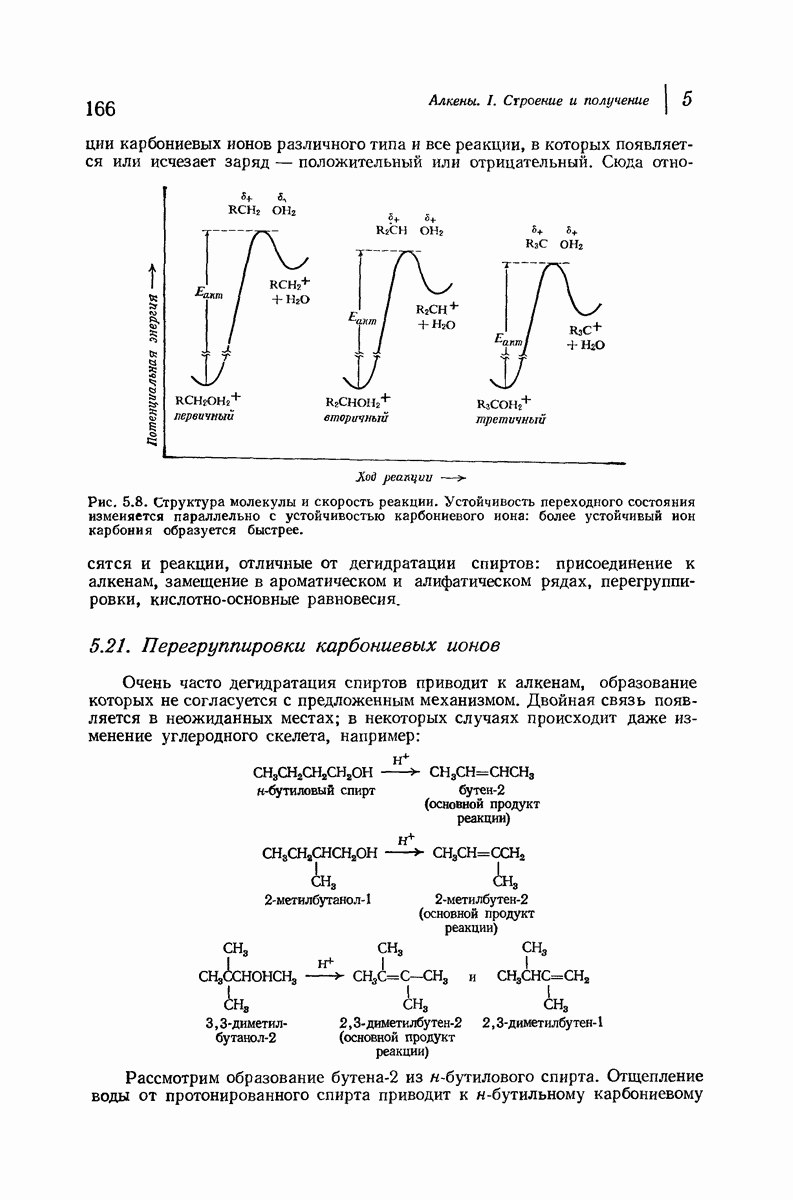
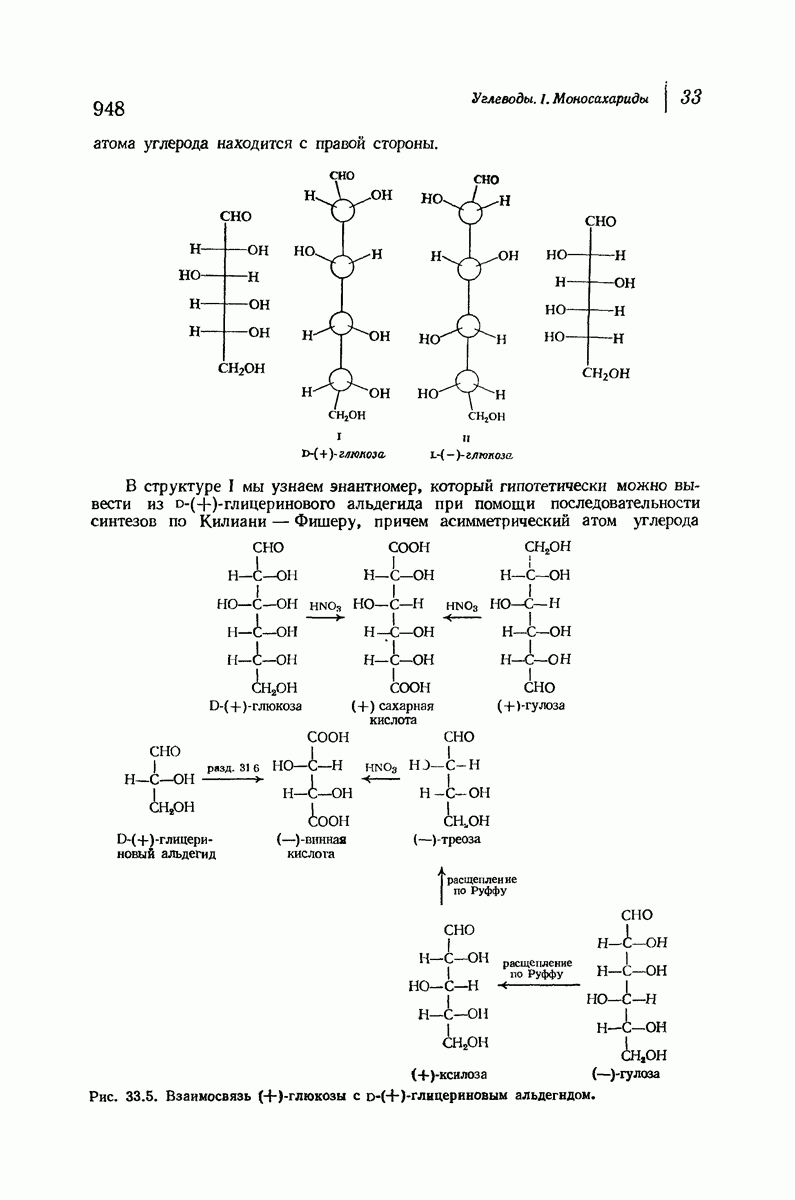
Прогресс, достигнутый в ходе решения столь сложный проблемы, был, естественно, результатом усилий многих исследователей. Среди них — Лайнус Полинг (Калифорнийский технологический институт), получивший в 1954 г. Нобелевскую премию. В 1951 г. Полинг писал: «Четырнадцать лет назад профессор Р. Кори и я, предприняв очень энергичные, но безуспешные попытки решить задачу построения удовлетворительной модели конфигурации полипептидных цепей в белках, решили попытаться справиться с этой задачей косвенным методом, тщательно изучив кристаллы аминокислот, простых пептидов и родственных соединений для того, чтобы получить абсолютно надежные и подробные сведения о структурных характеристиках веществ подобного рода и в конце концов получить возможность уверенного предсказания точных конфигураций полипептидных цепей в белках» [Record. Chem. Prog., 12, 156—157 (1951)]. Эта работа на простых веществах, проводившаяся в течение более 14 лет, позволила в конце концов Полингу с сотрудниками предложить структуру, которая, вероятно, является важнейшей вторичной структурой в химии белков —  -спираль.
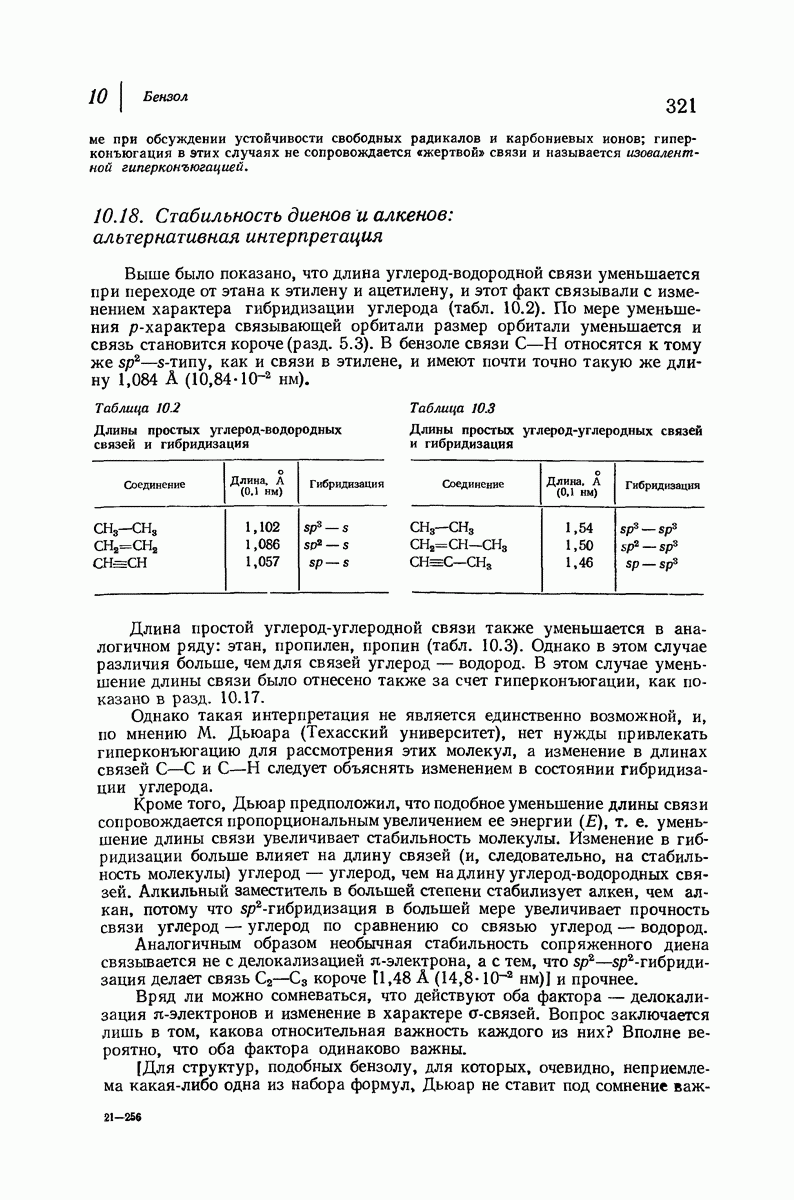
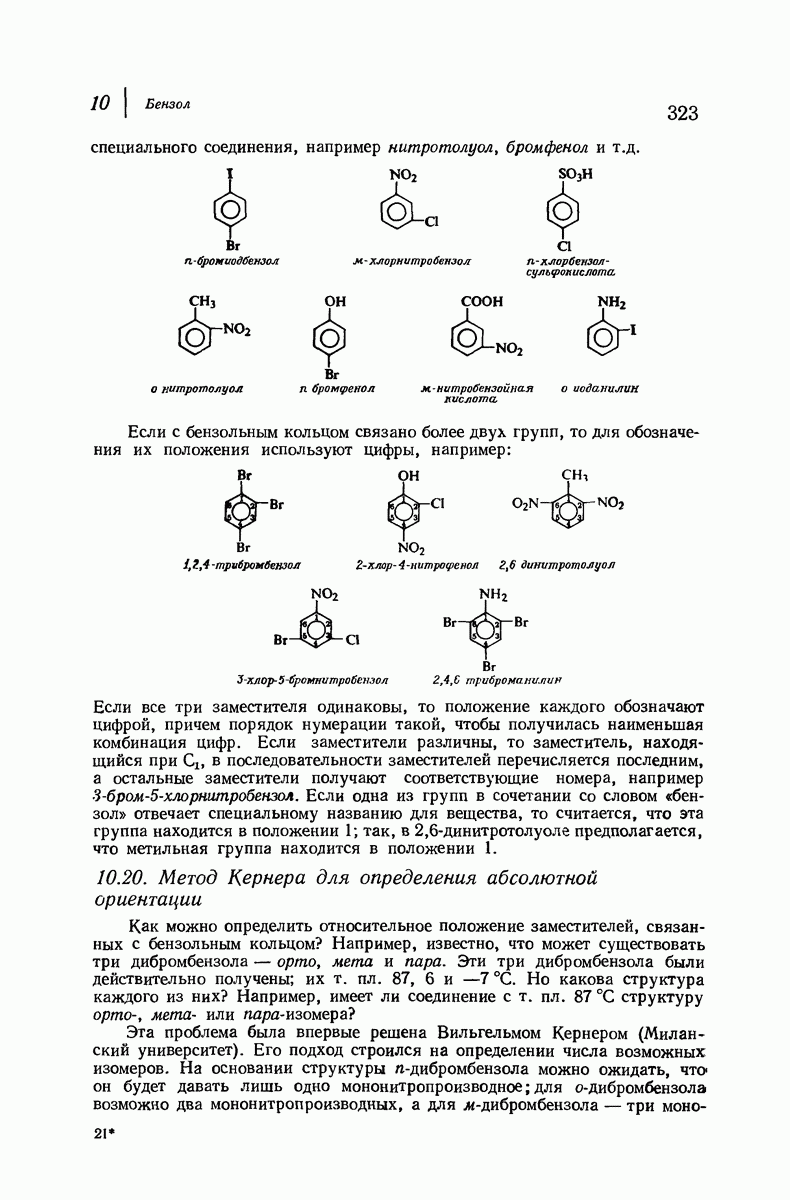
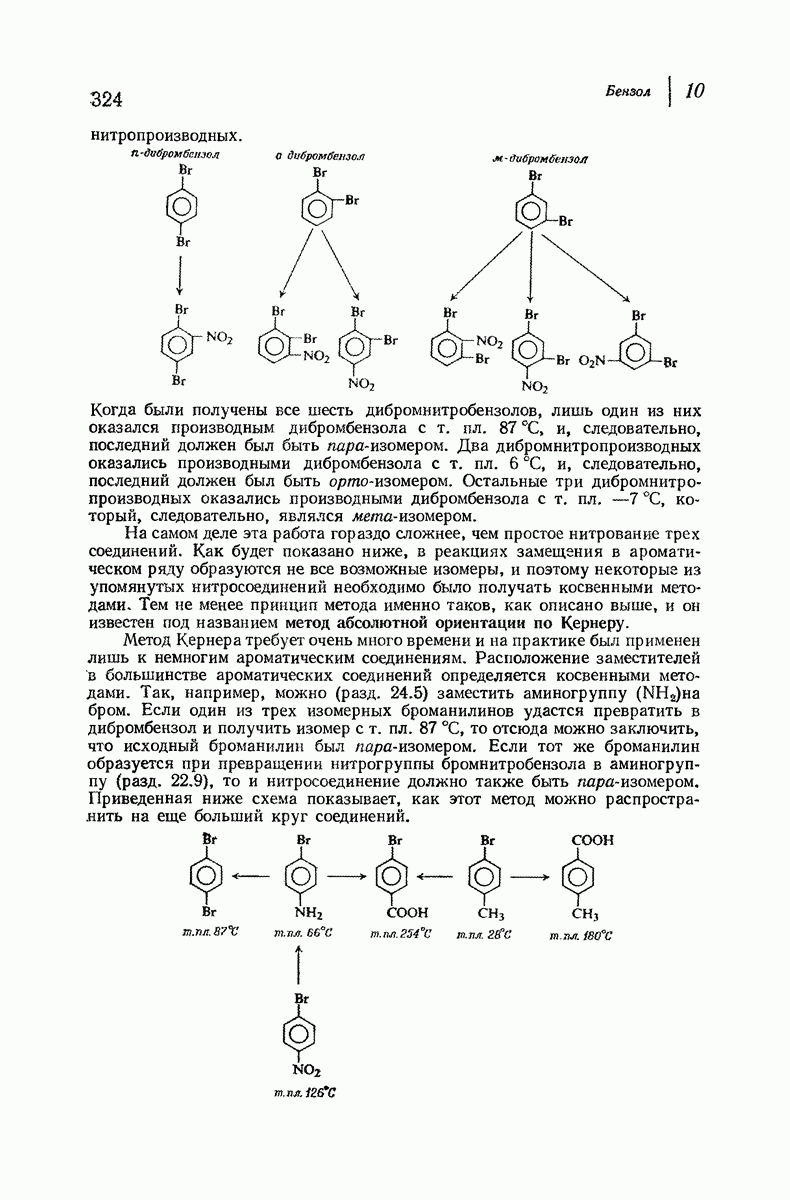
-спираль.
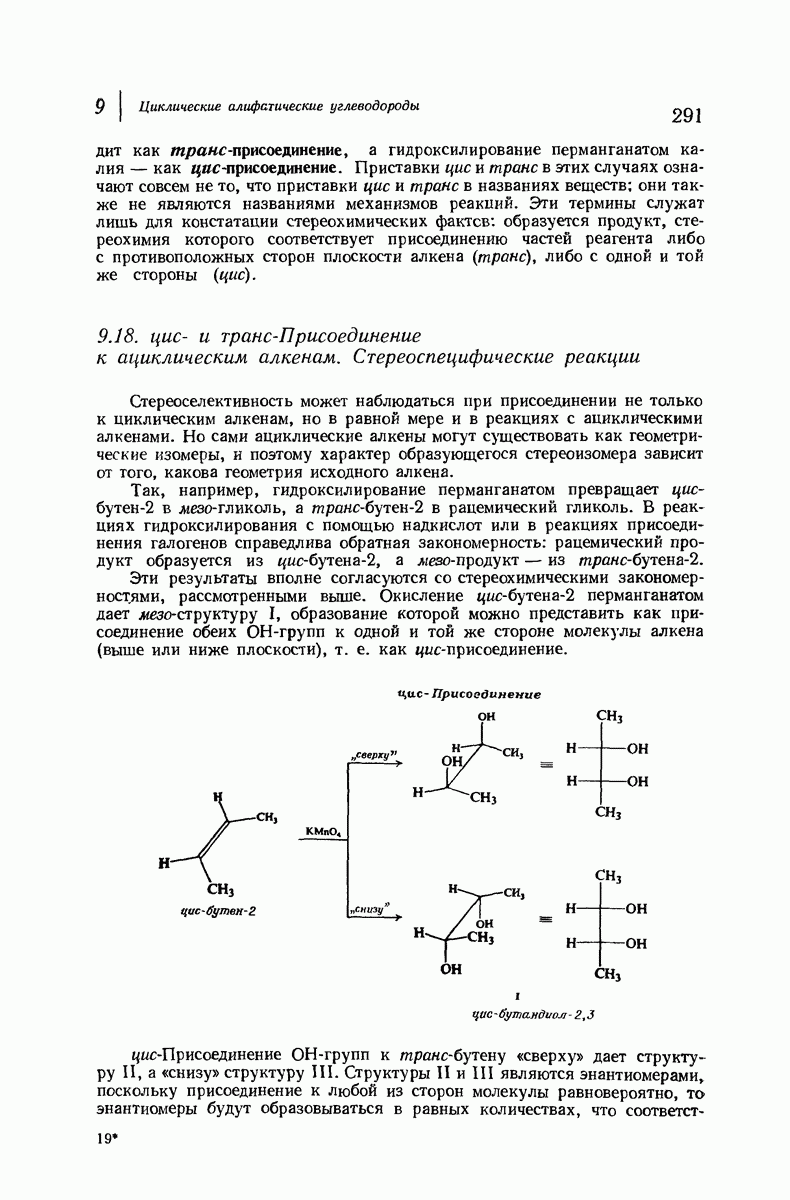
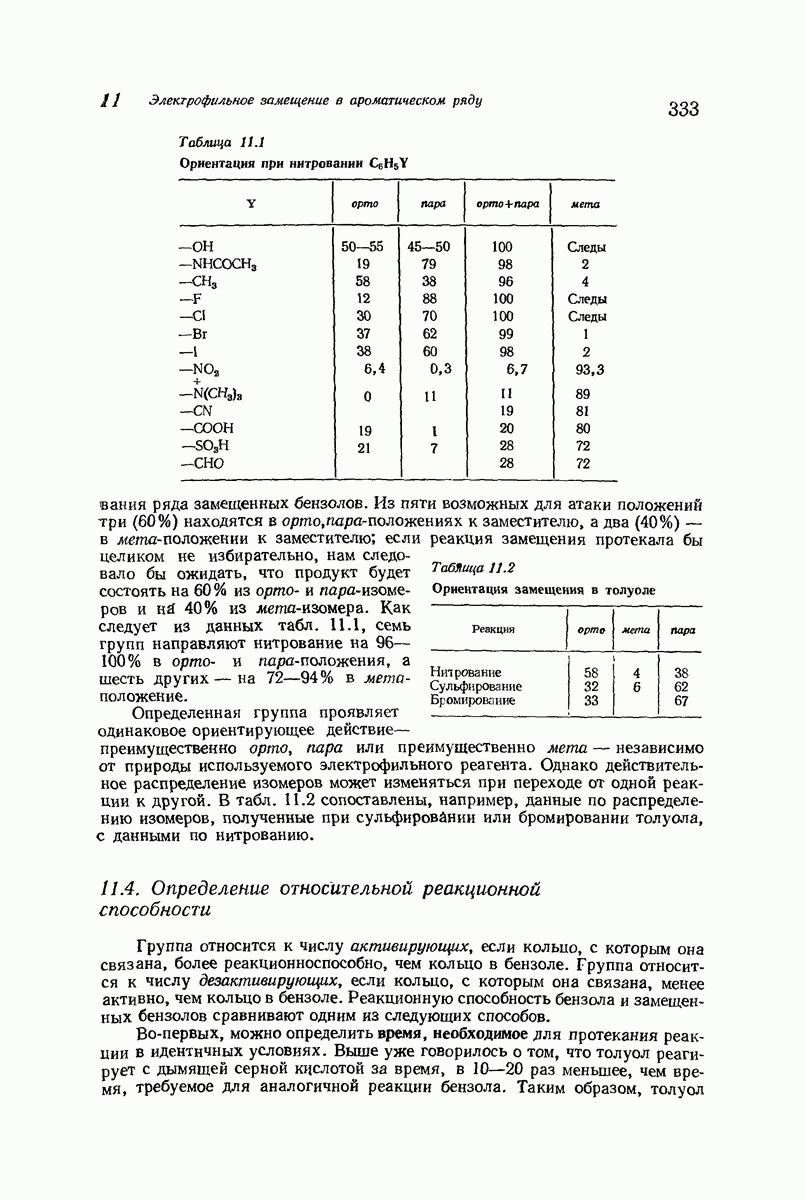
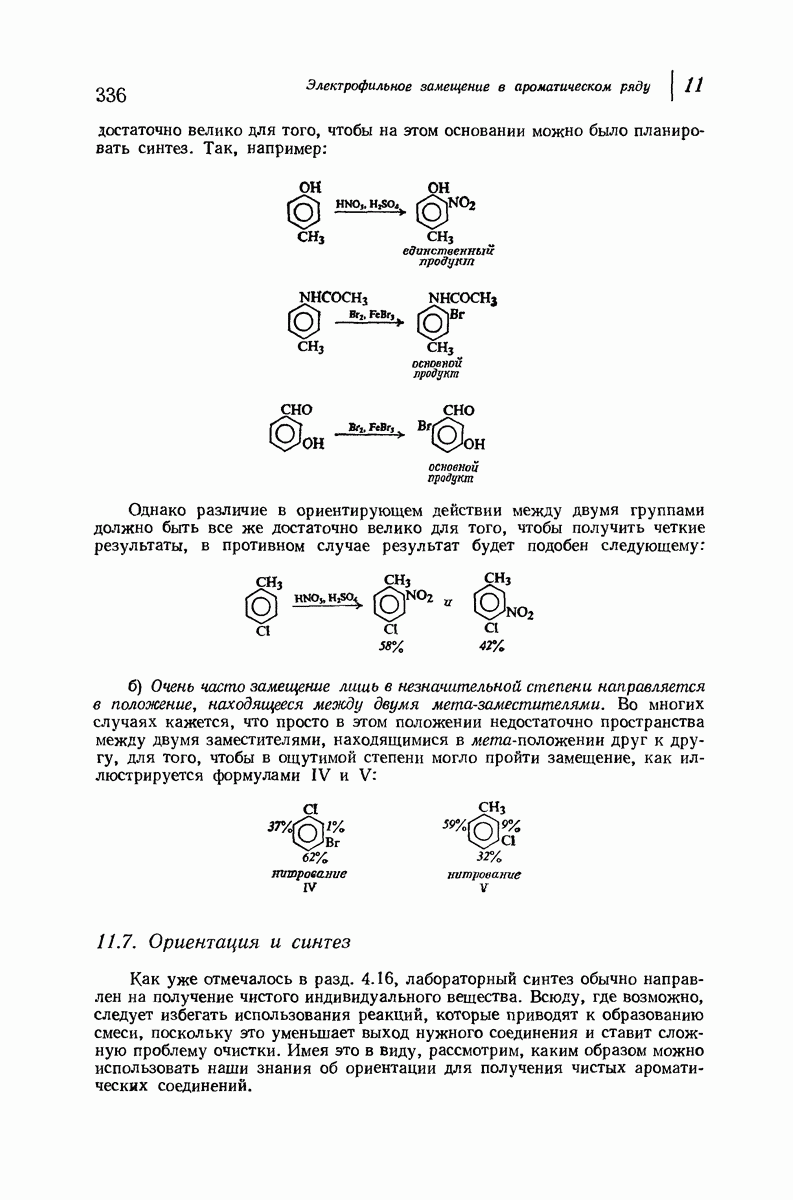
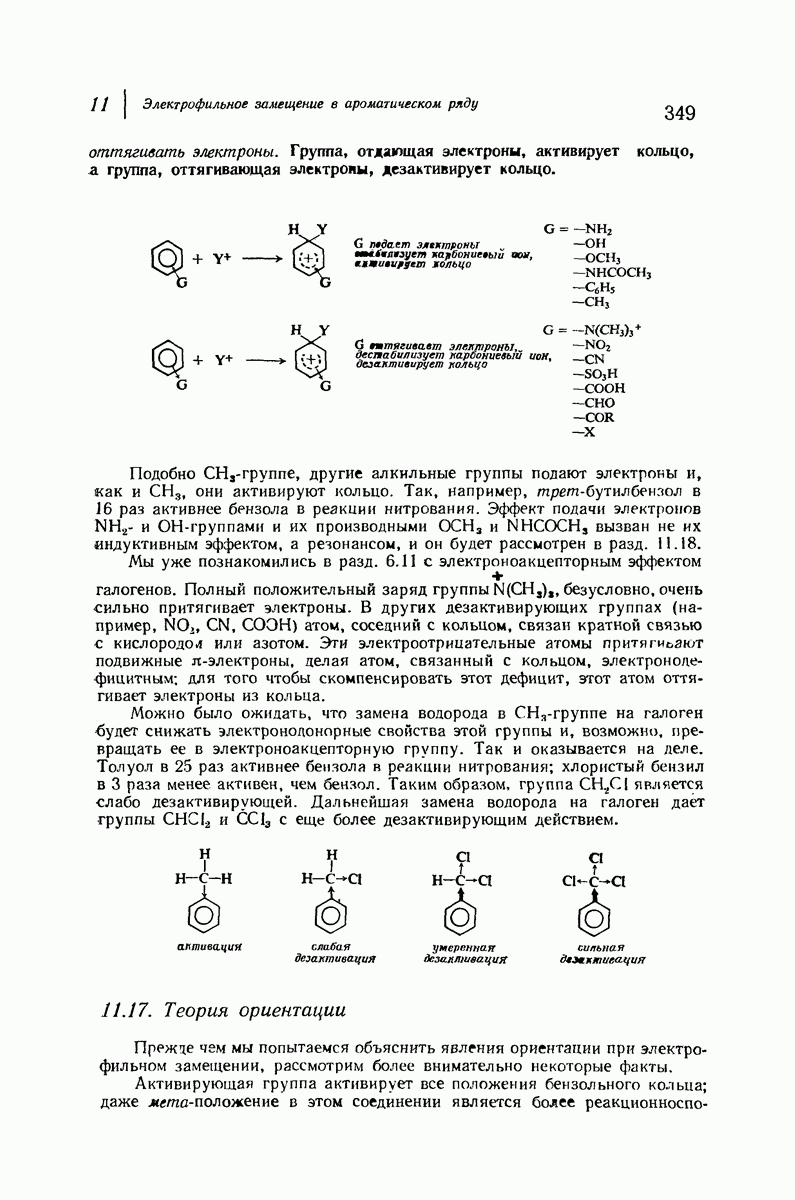
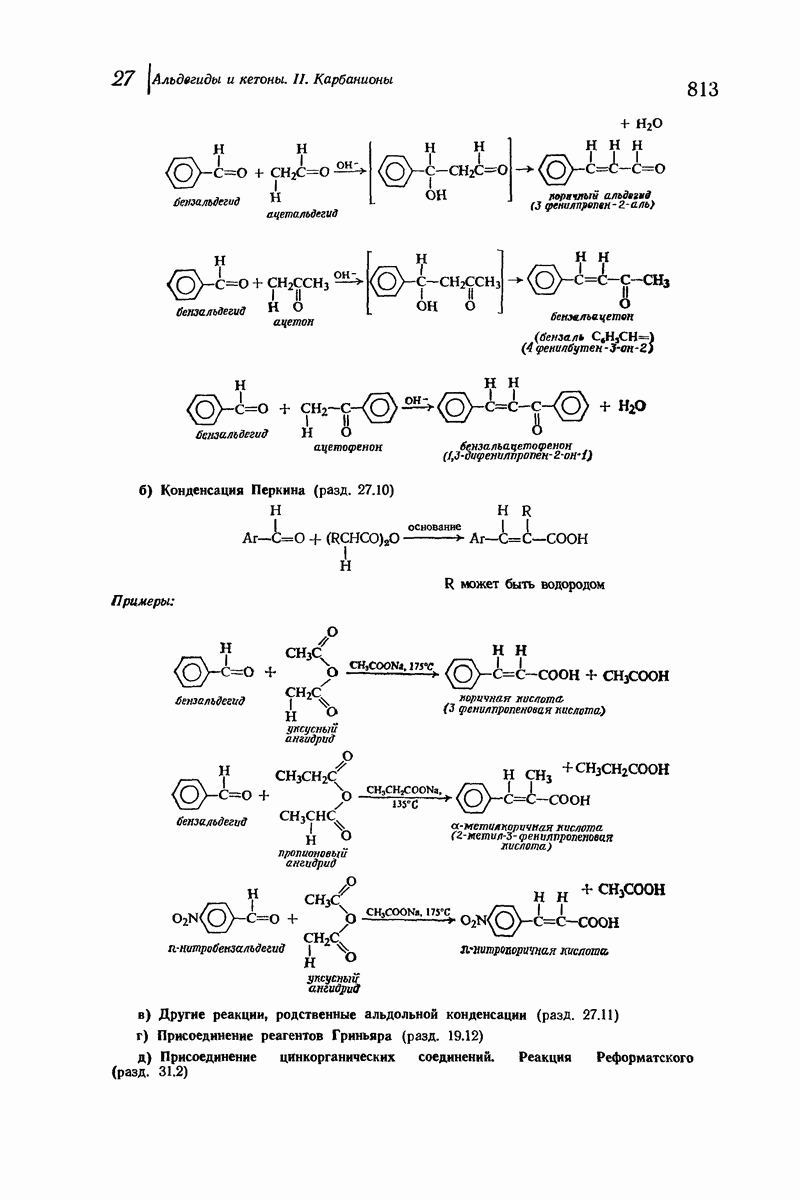
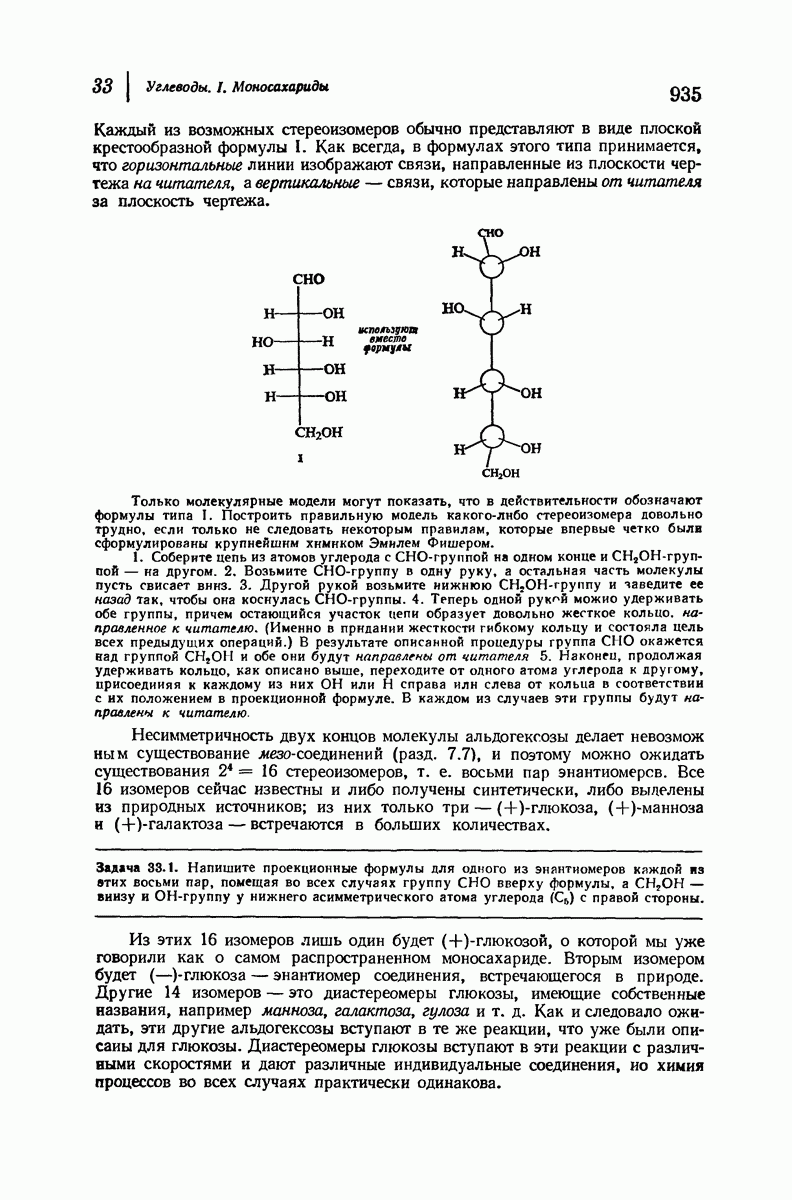
Рассмотрим некоторые из предложенных вторичных структур.
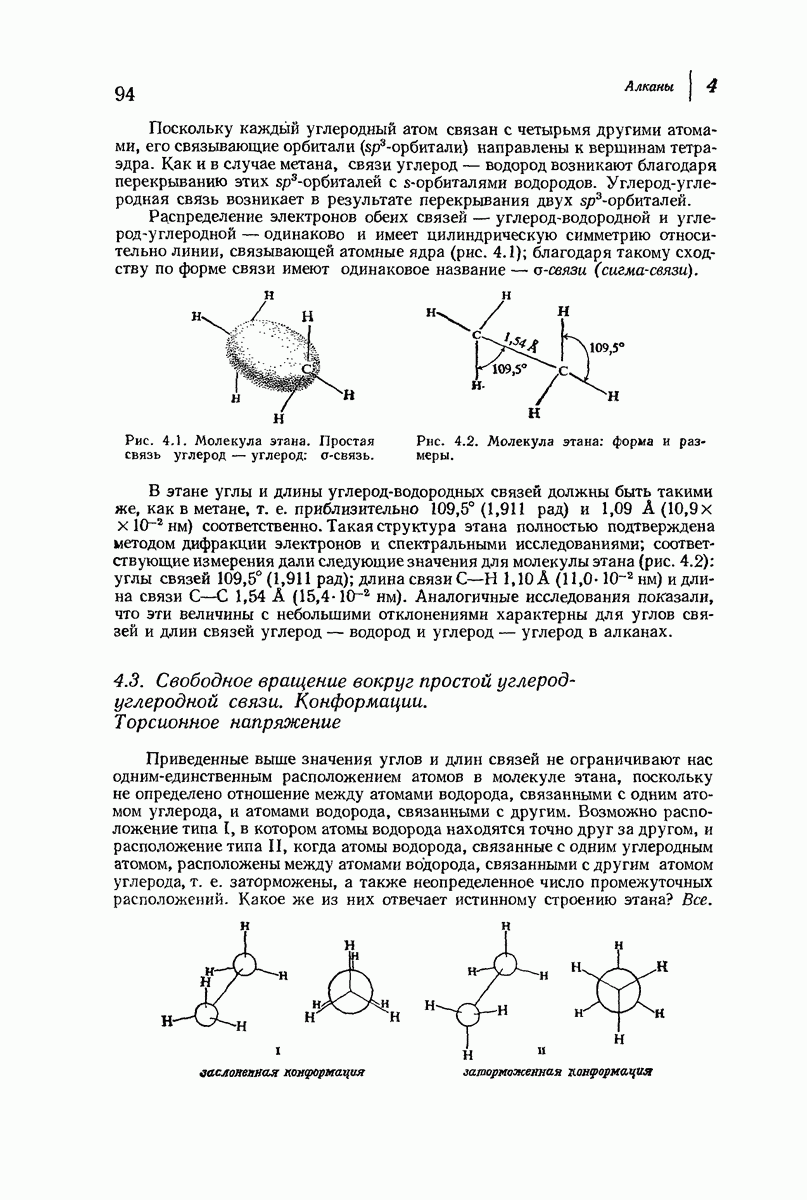
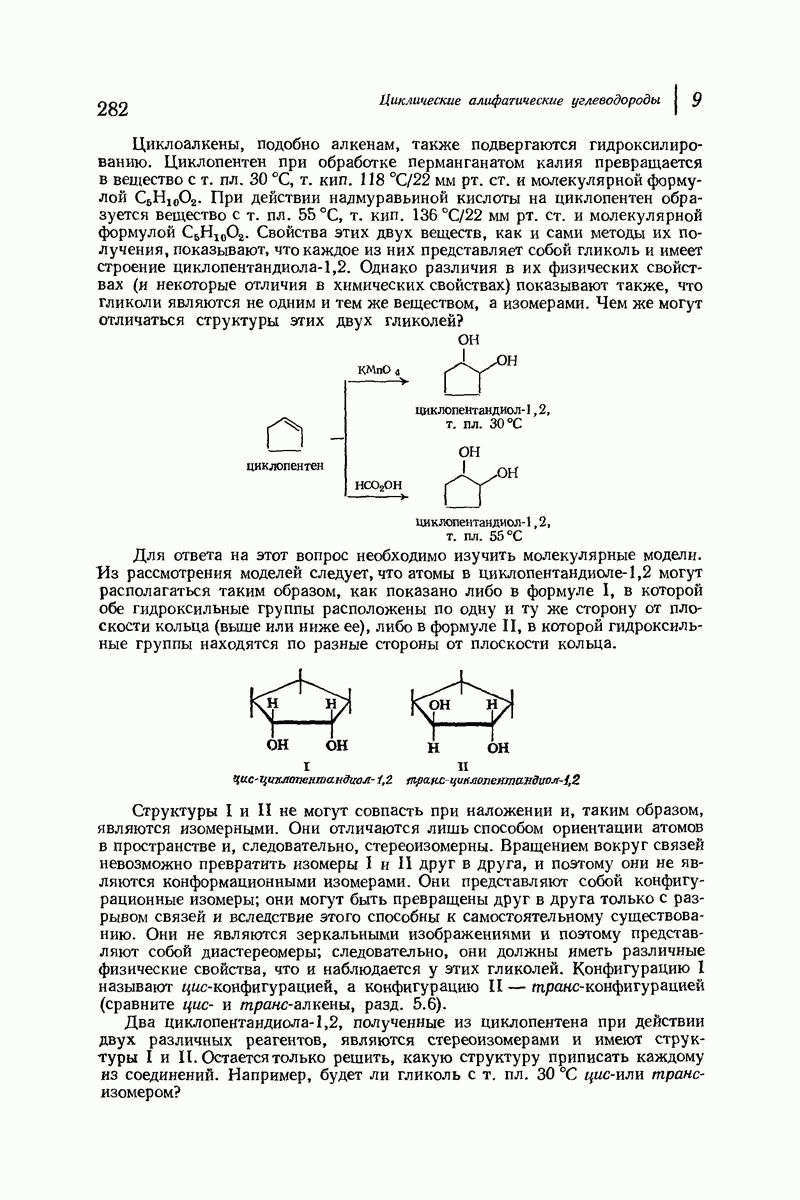
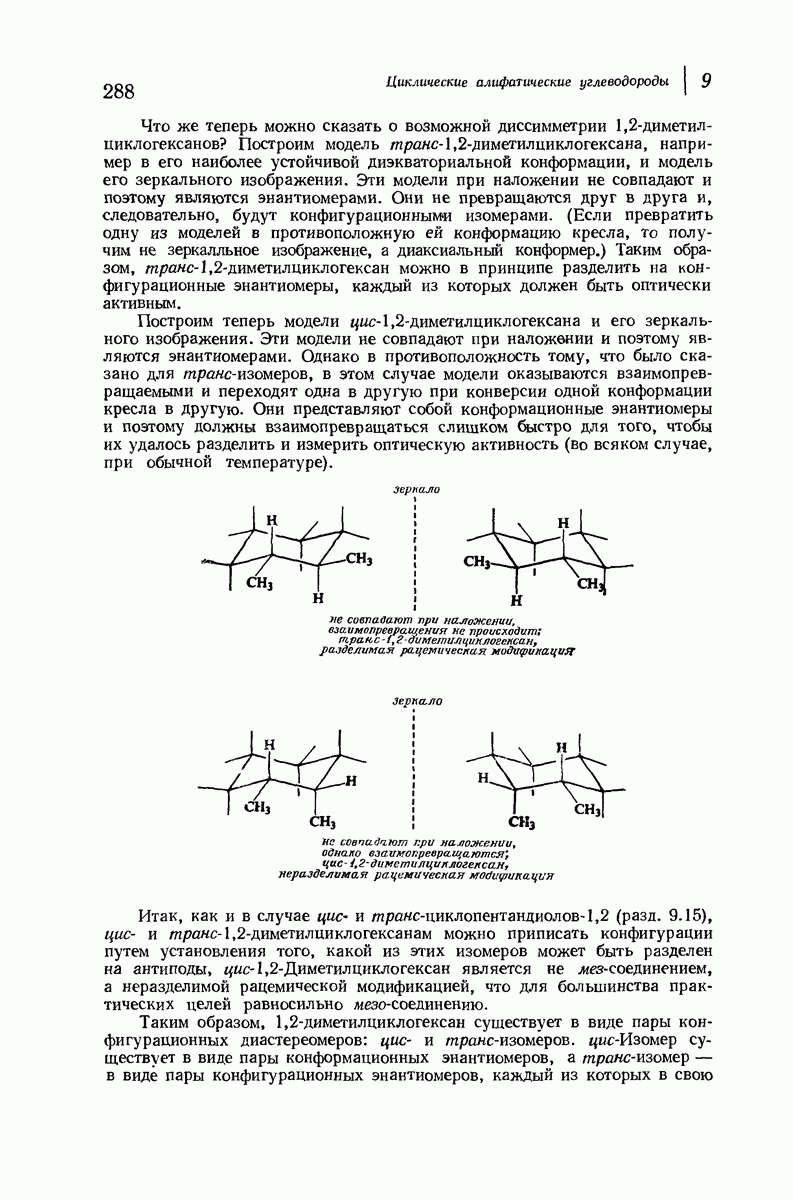
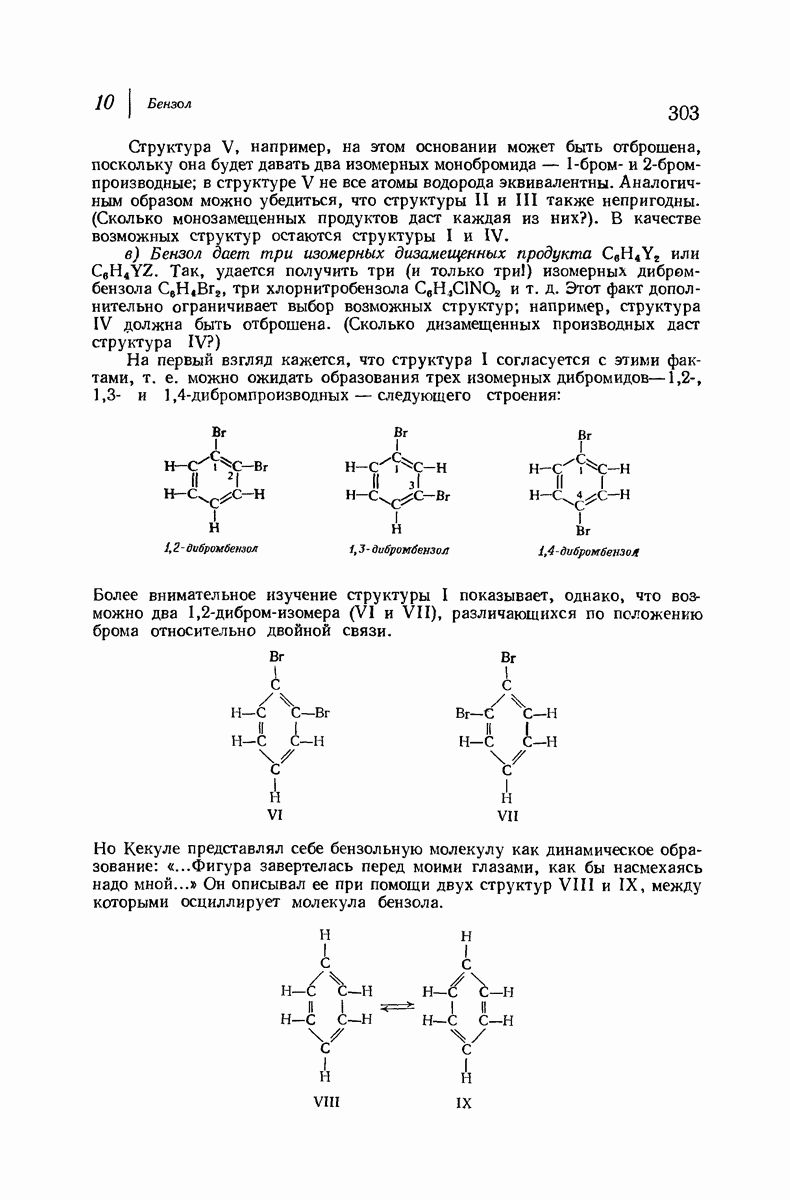
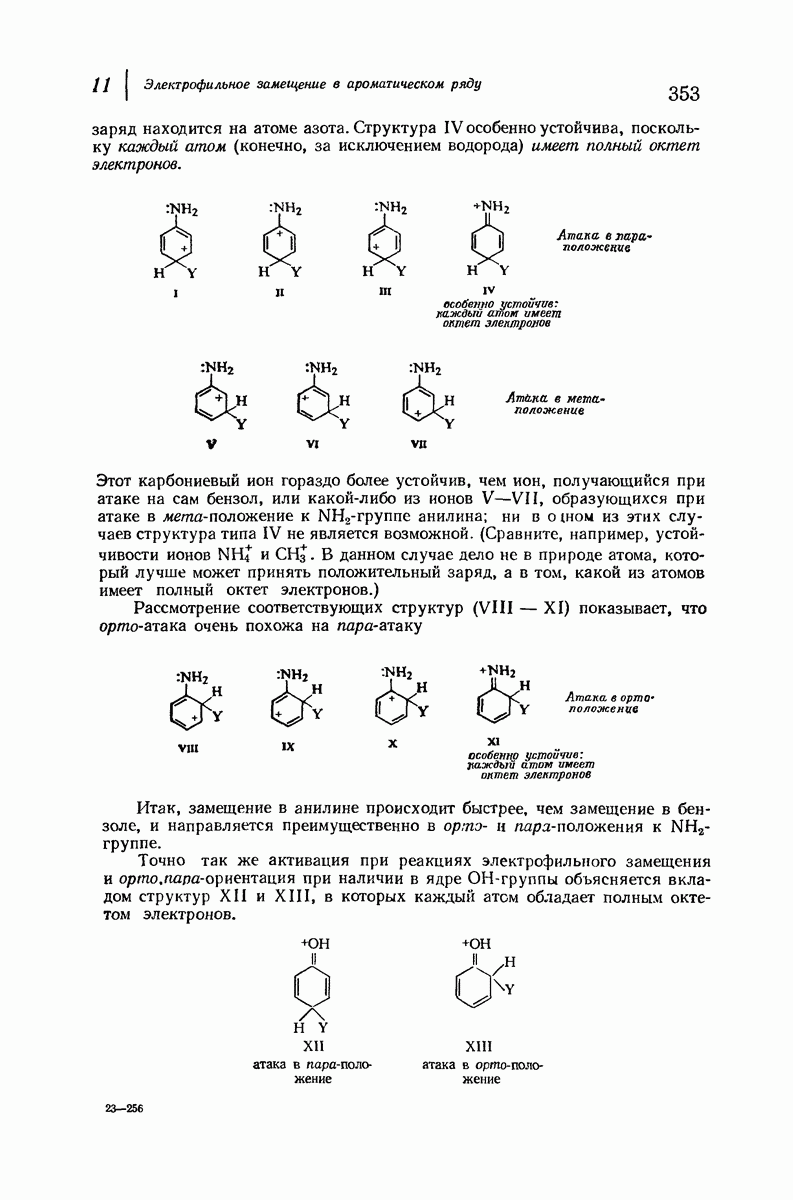
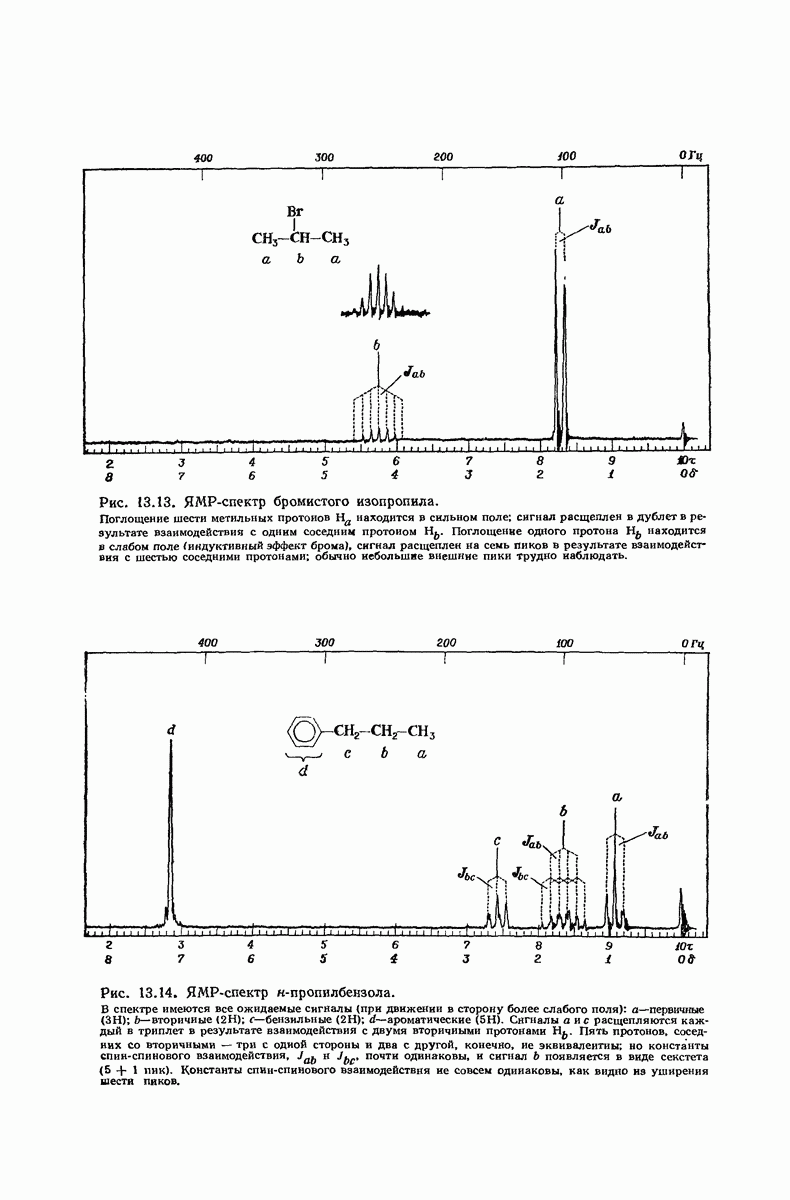
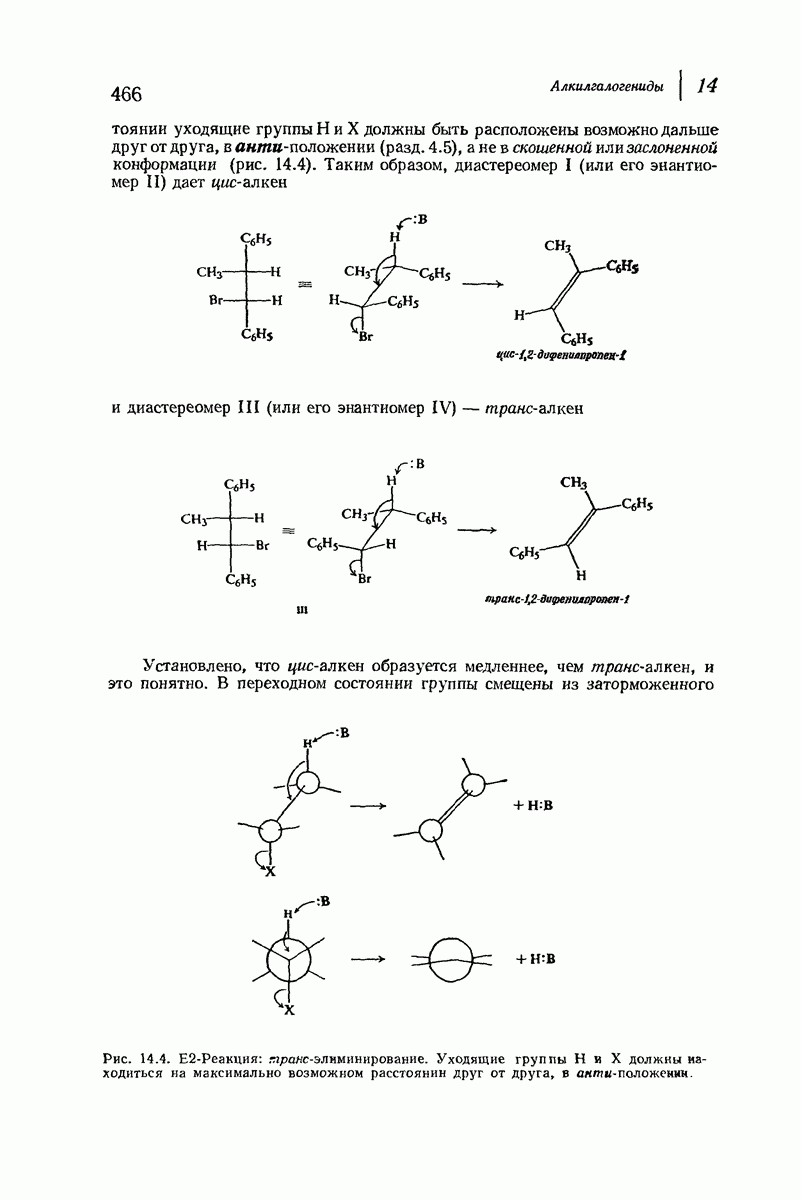
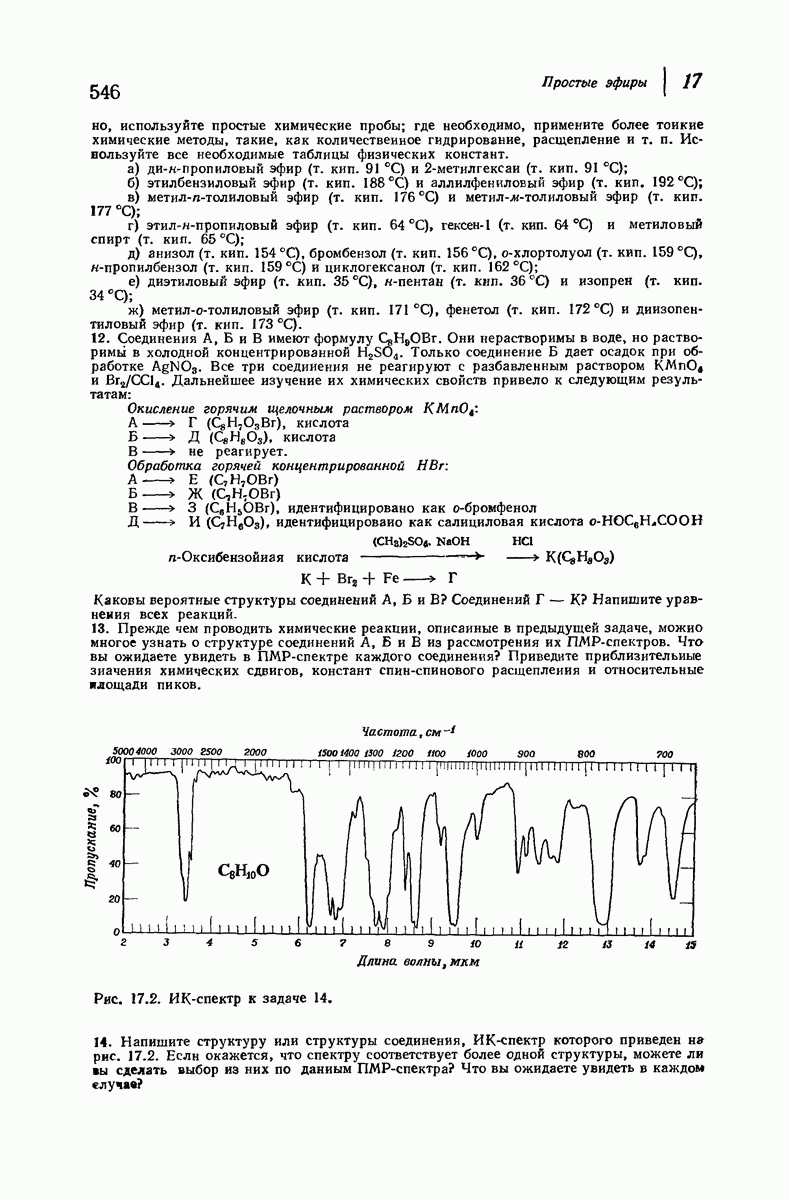
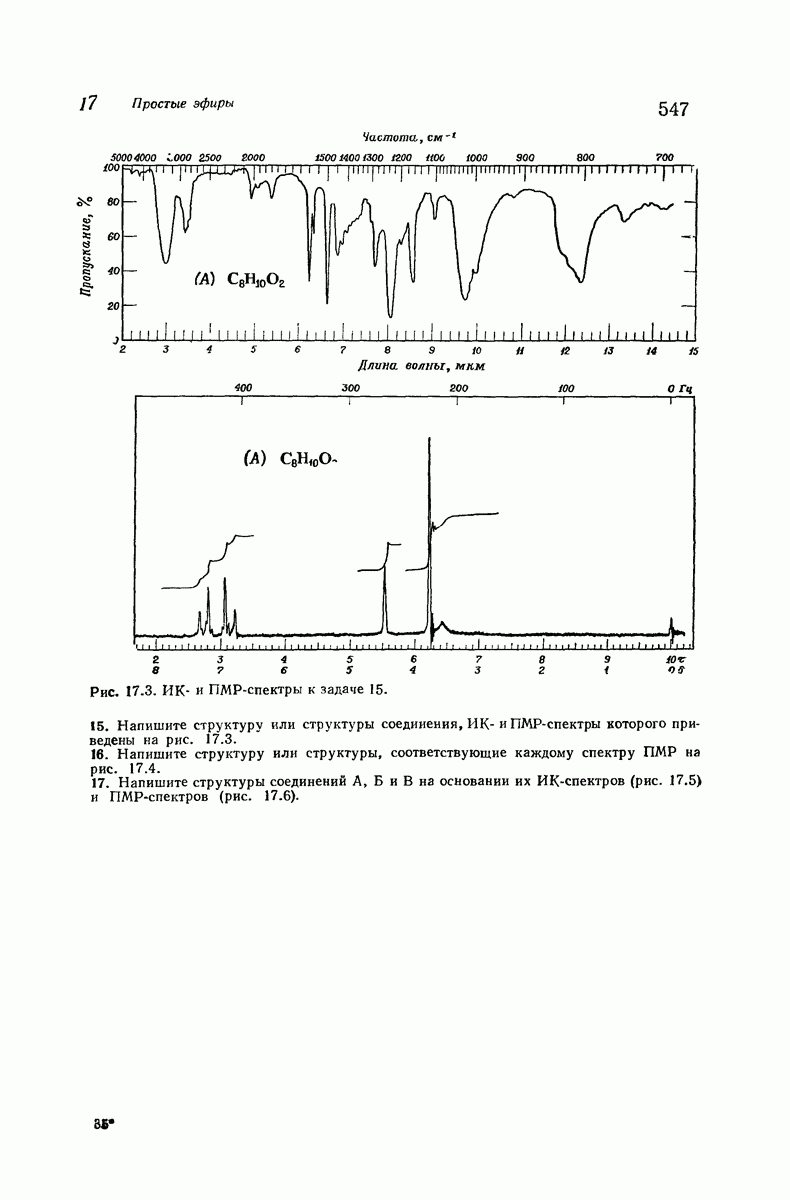
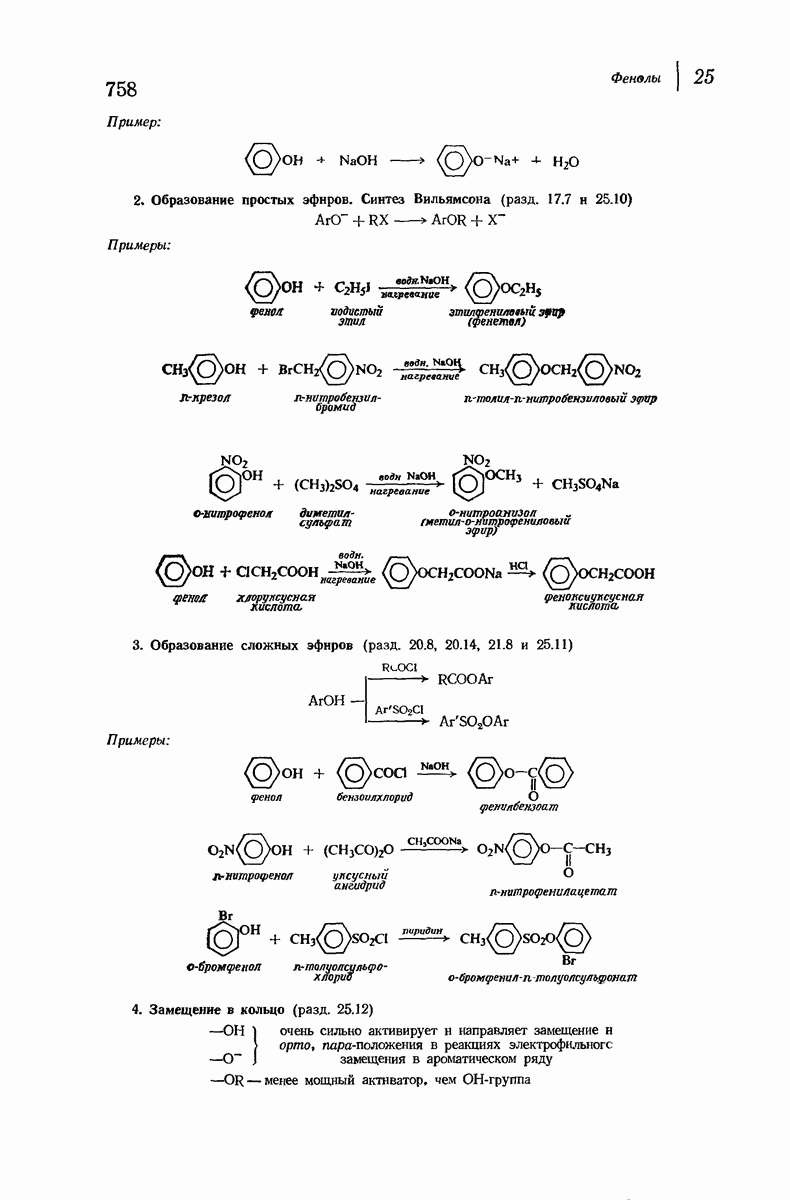
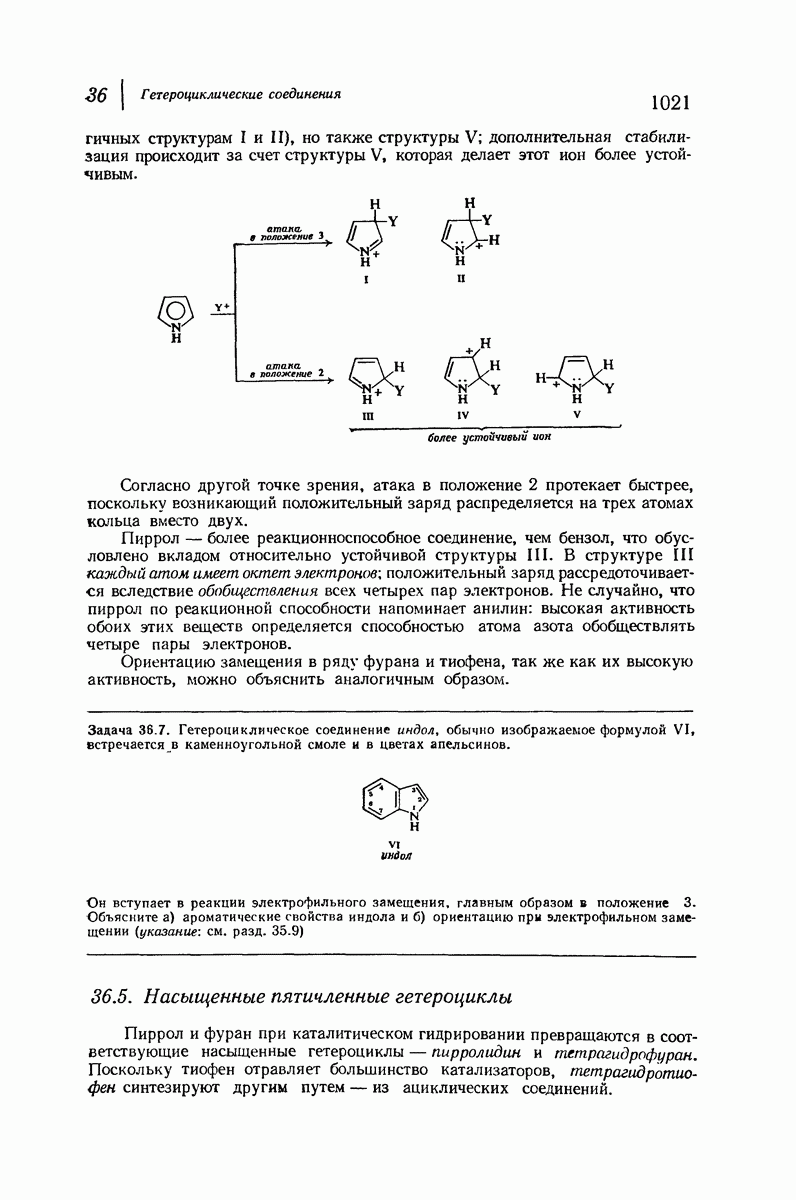
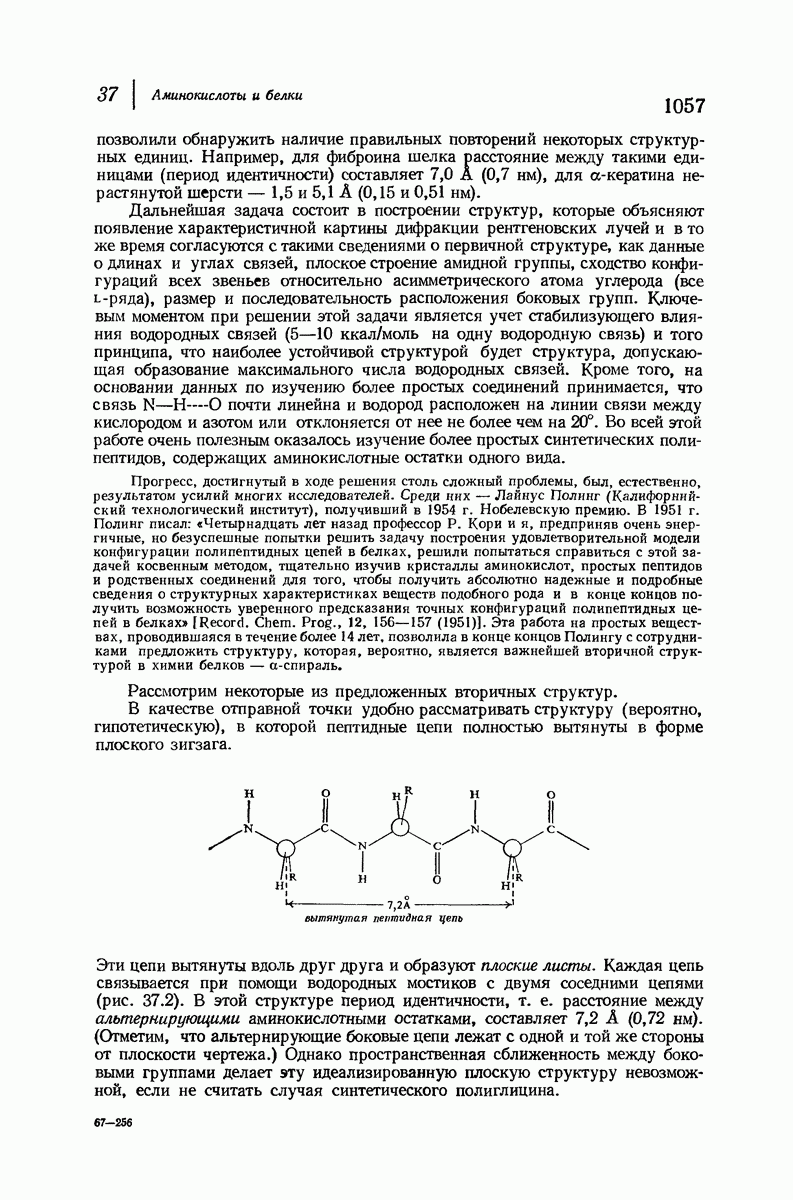
В качестве отправной точки удобно рассматривать структуру (вероятно, гипотетическую), в которой пептидные цепи полностью вытянуты в форме плоского зигзага.

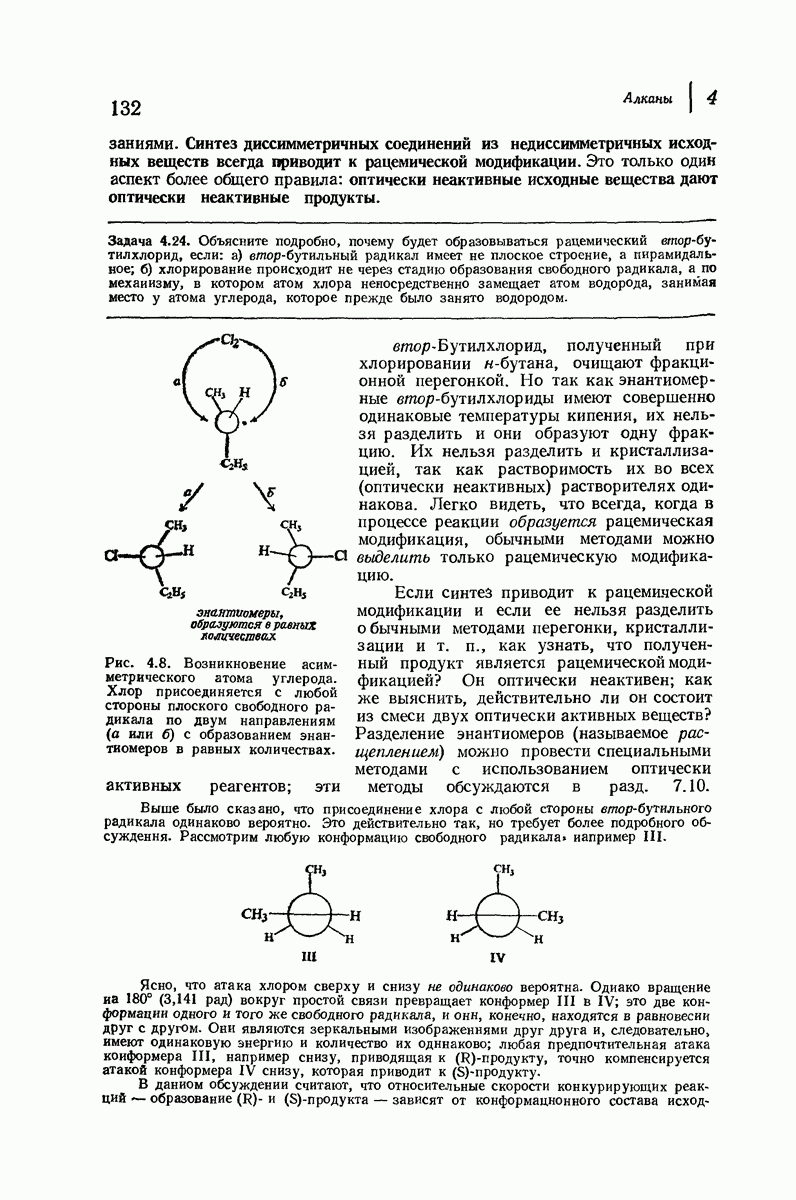
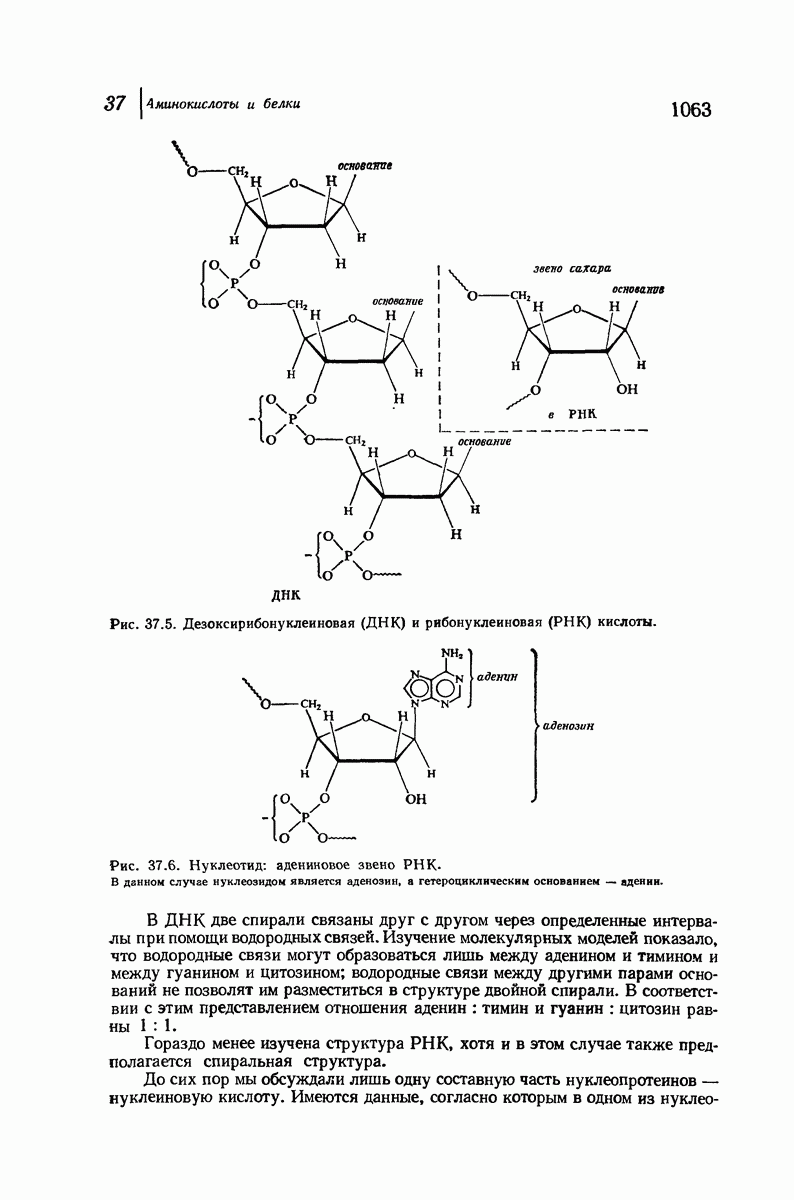
Эти цепи вытянуты вдоль друг друга и образуют плоские листы. Каждая цепь связывается при помощи водородных мостиков с двумя соседними цепями (рис. 37.2). В этой структуре период идентичности, т. е. расстояние между альтернирующими аминокислотными остатками, составляет 7,2 А  (Отметим, что альтернирующие боковые цепи лежат с одной и той же стороны от плоскости чертежа.) Однако пространственная сближенность между боковыми группами делает эту идеализированную плоскую структуру невозможной, если не считать случая синтетического полиглицина.
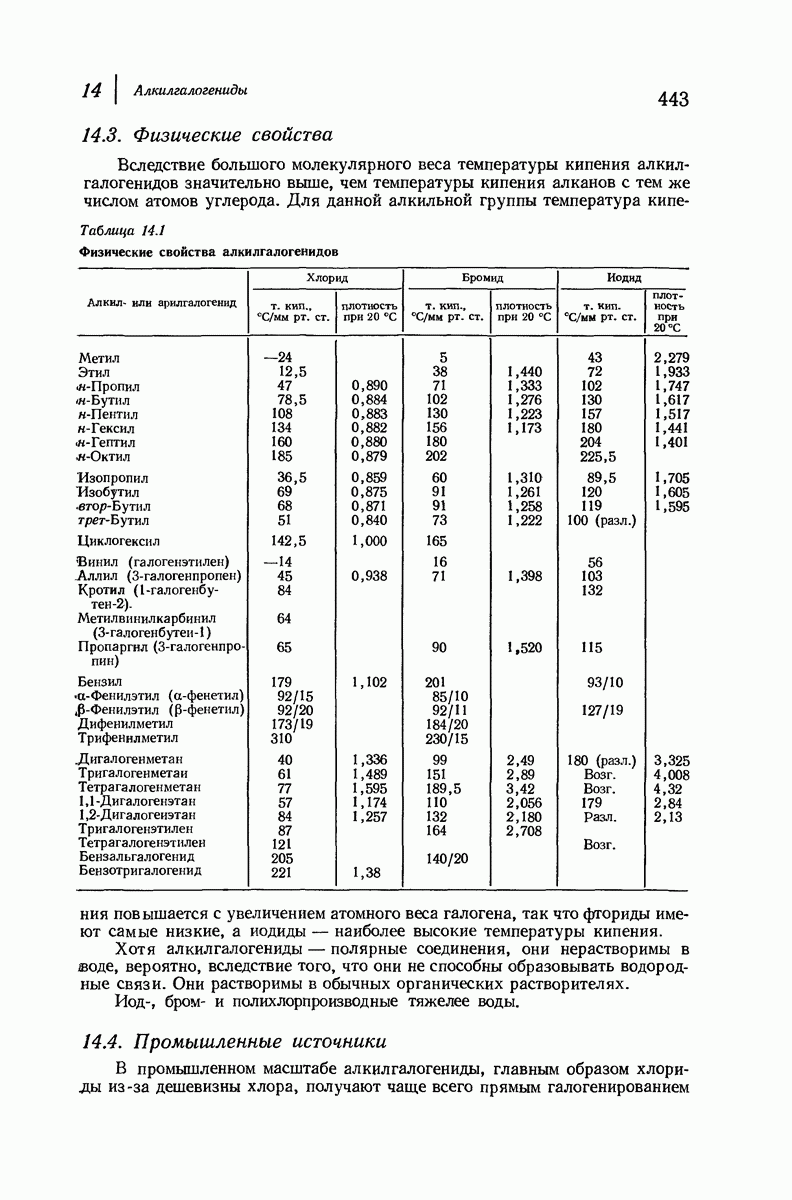
(Отметим, что альтернирующие боковые цепи лежат с одной и той же стороны от плоскости чертежа.) Однако пространственная сближенность между боковыми группами делает эту идеализированную плоскую структуру невозможной, если не считать случая синтетического полиглицина.
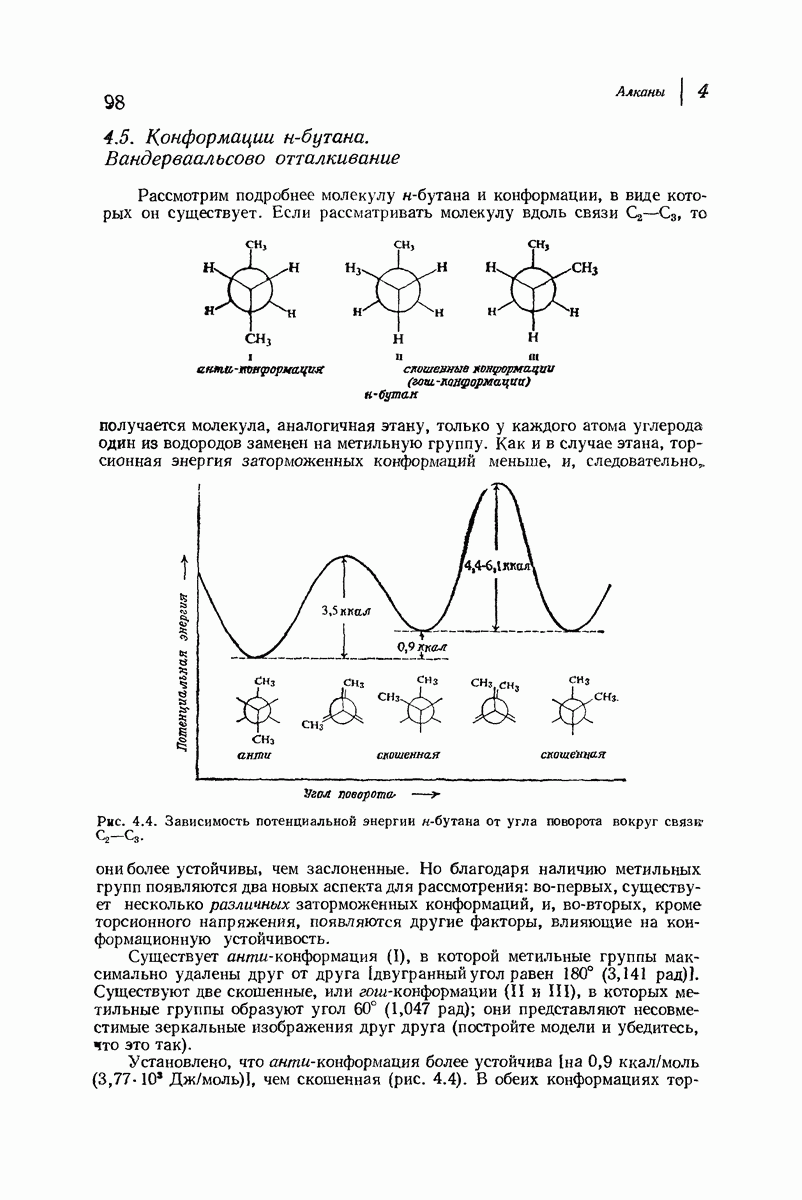
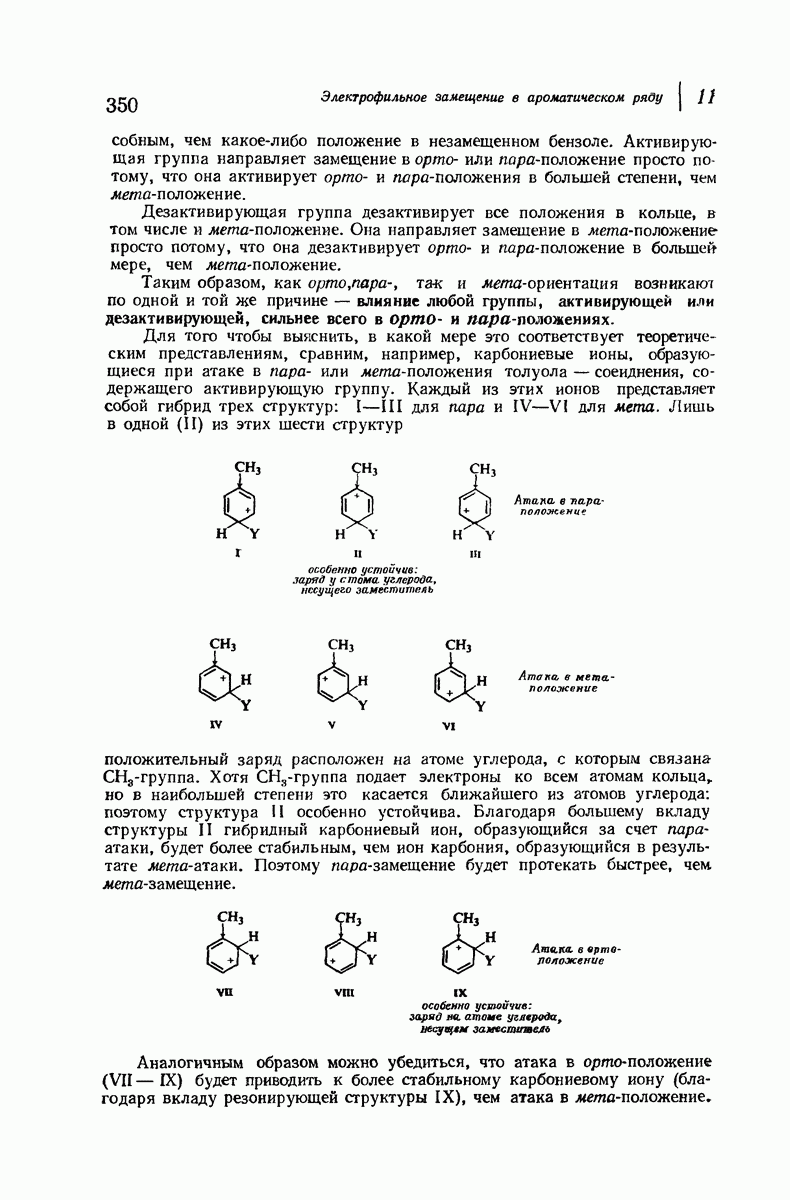
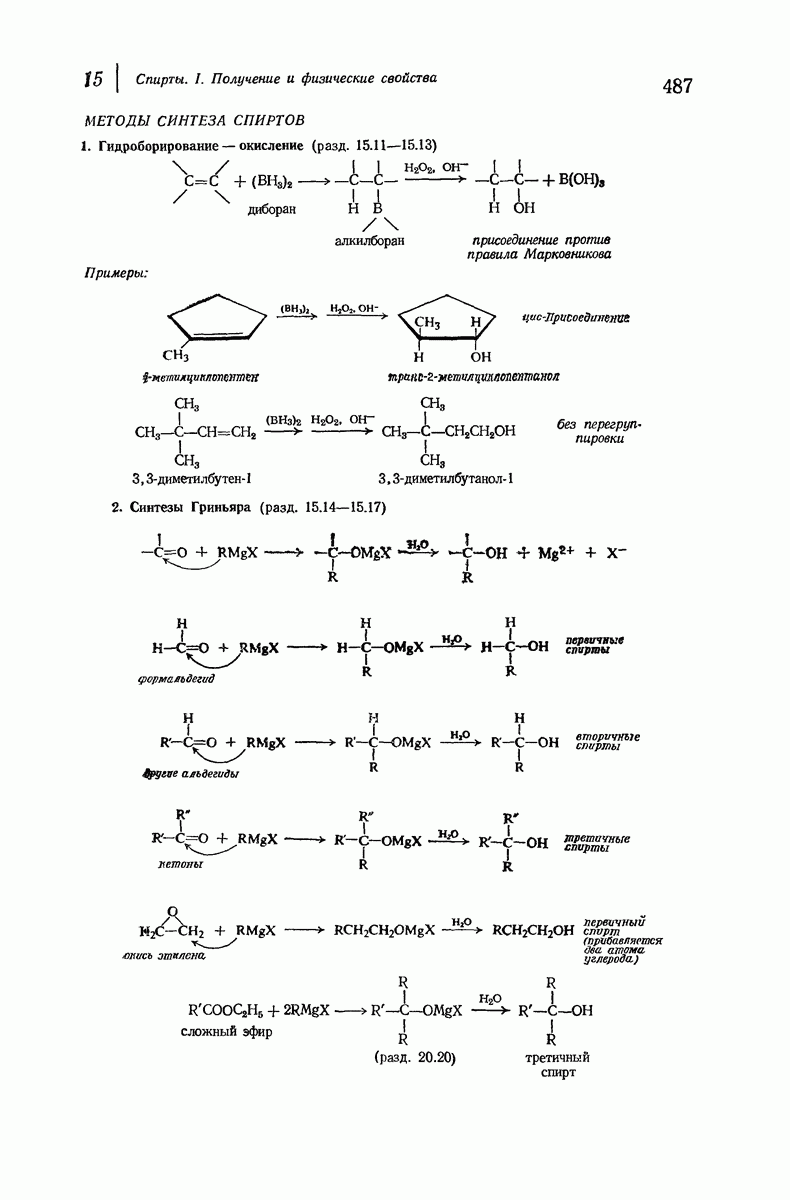
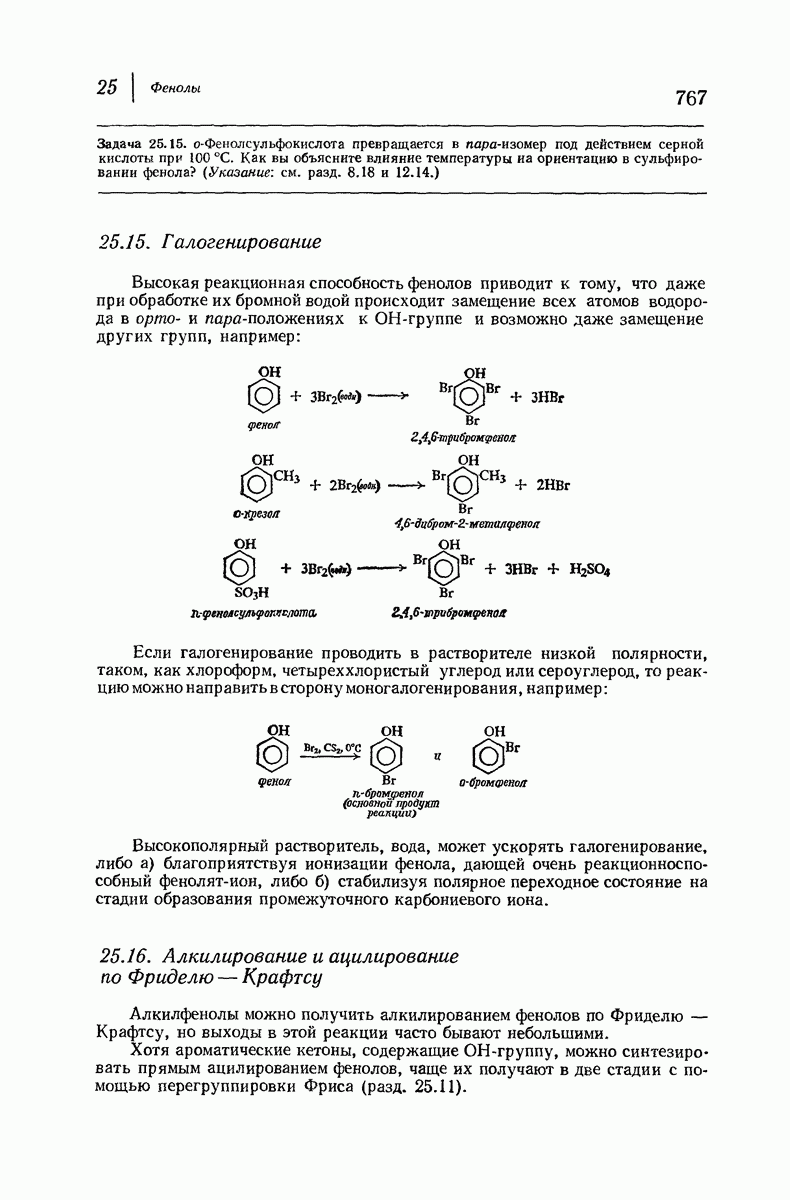
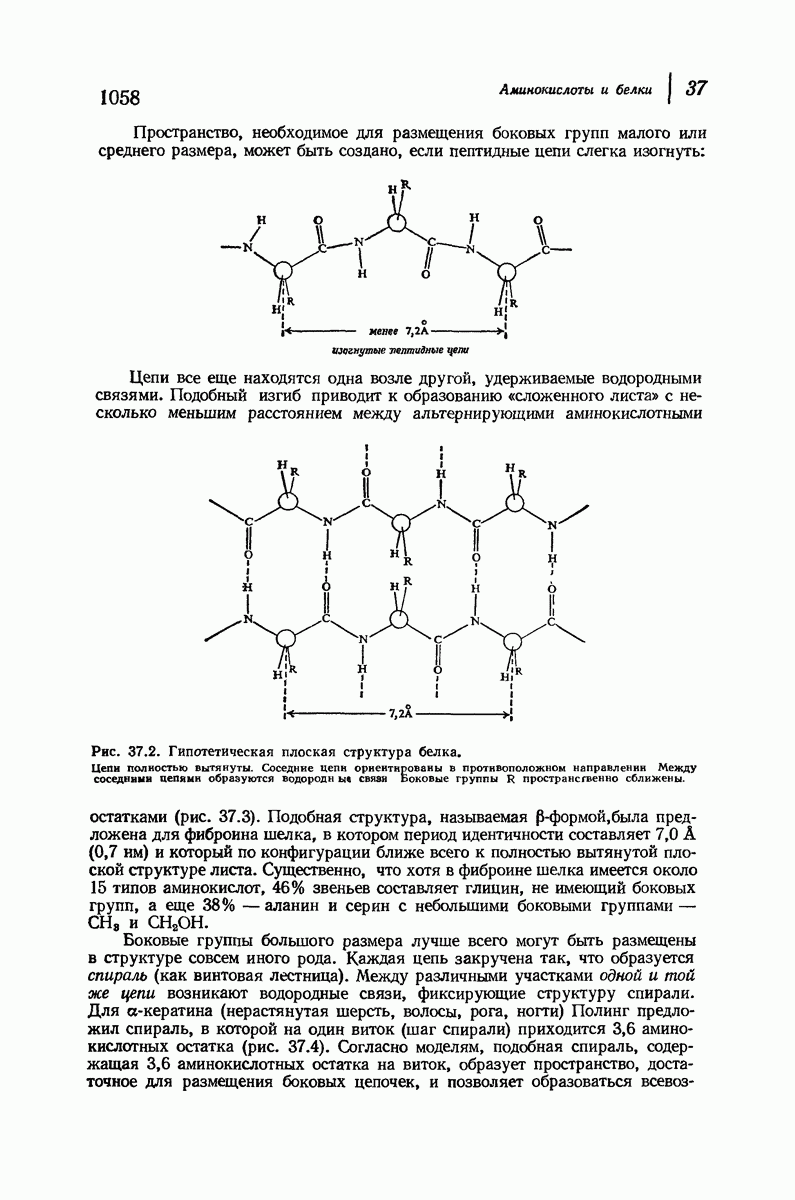
Пространство, необходимое для размещения боковых групп малого или среднего размера, может быть создано, если пептидные цепи слегка изогнуть:

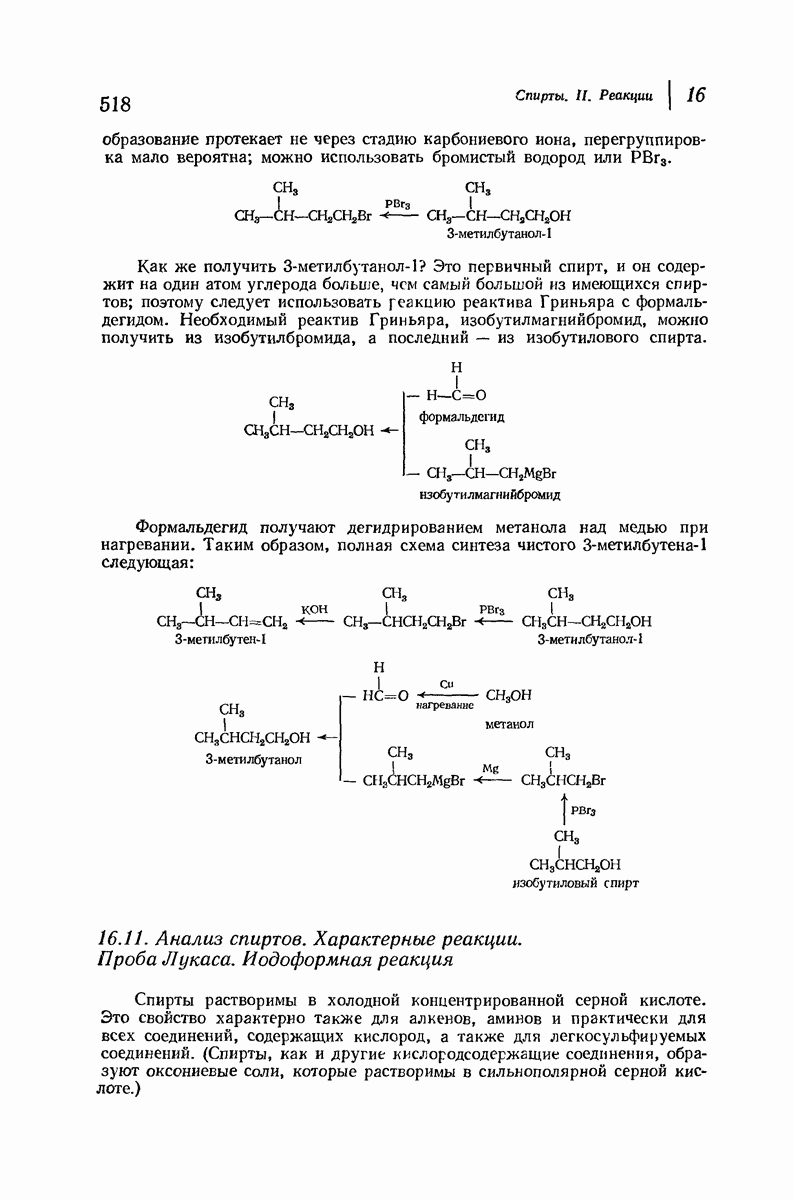
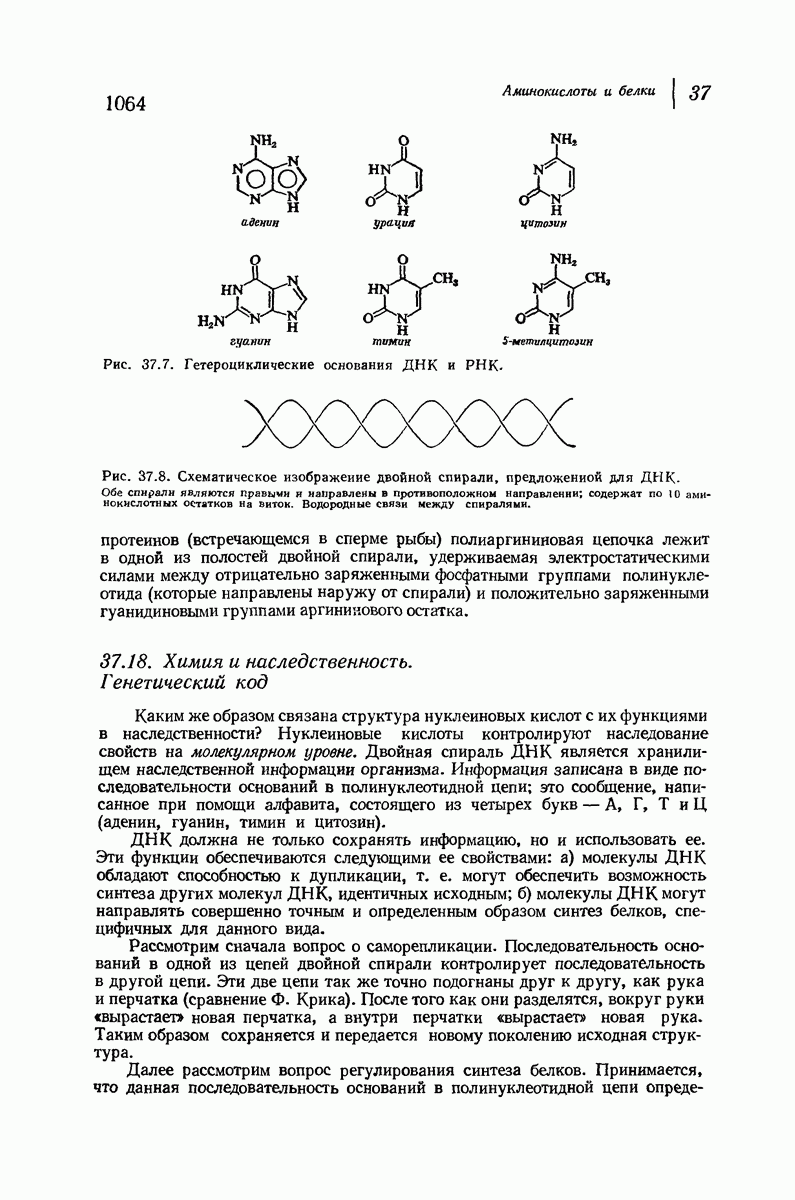
Цепи все еще находятся одна возле другой, удерживаемые водородными связями. Подобный изгиб приводит к образованию «сложенного листа» с несколько меньшим расстоянием между альтернирующими аминокислотными остатками (рис. 37.3).

Рис. 37.2. Гипотетическая плоская структура белка. Цепи полностью вытянуты. Соседние цепн ориентированы в противоположном направлении Между соседннын цепями образуются водородные связи. Боковые группы R пространственно сближены.
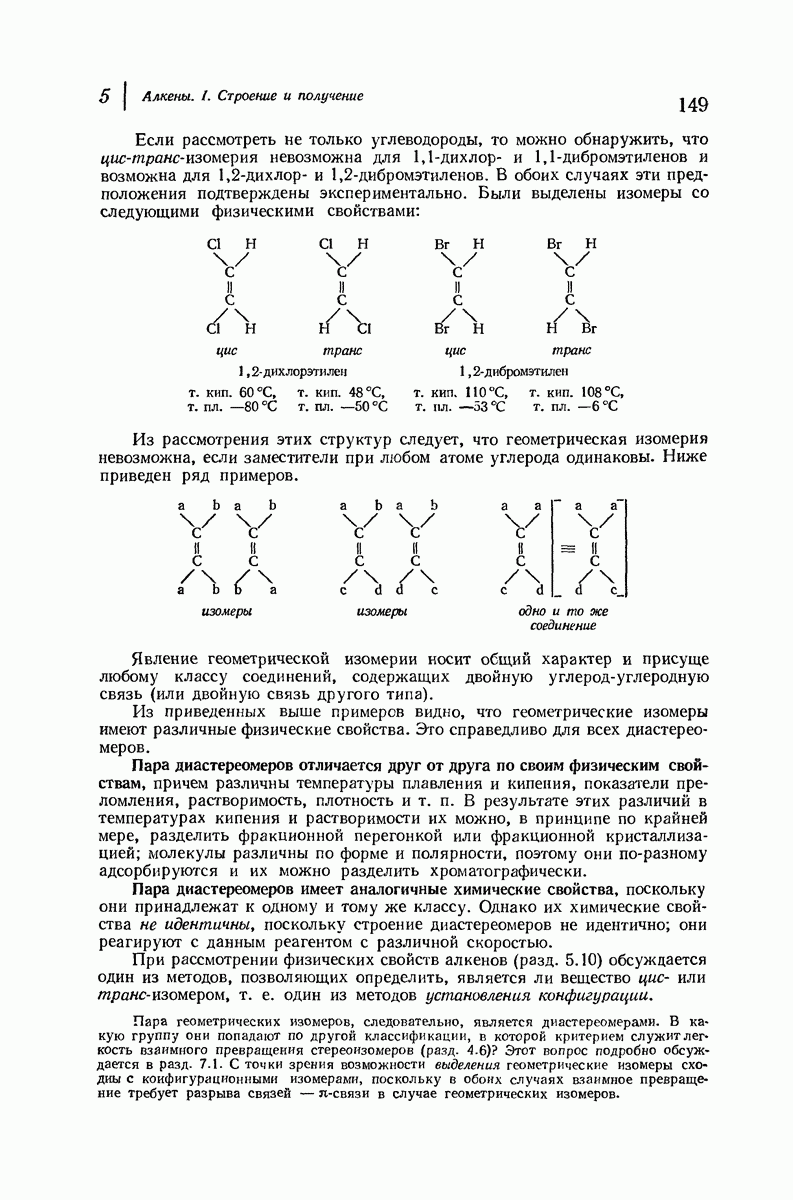
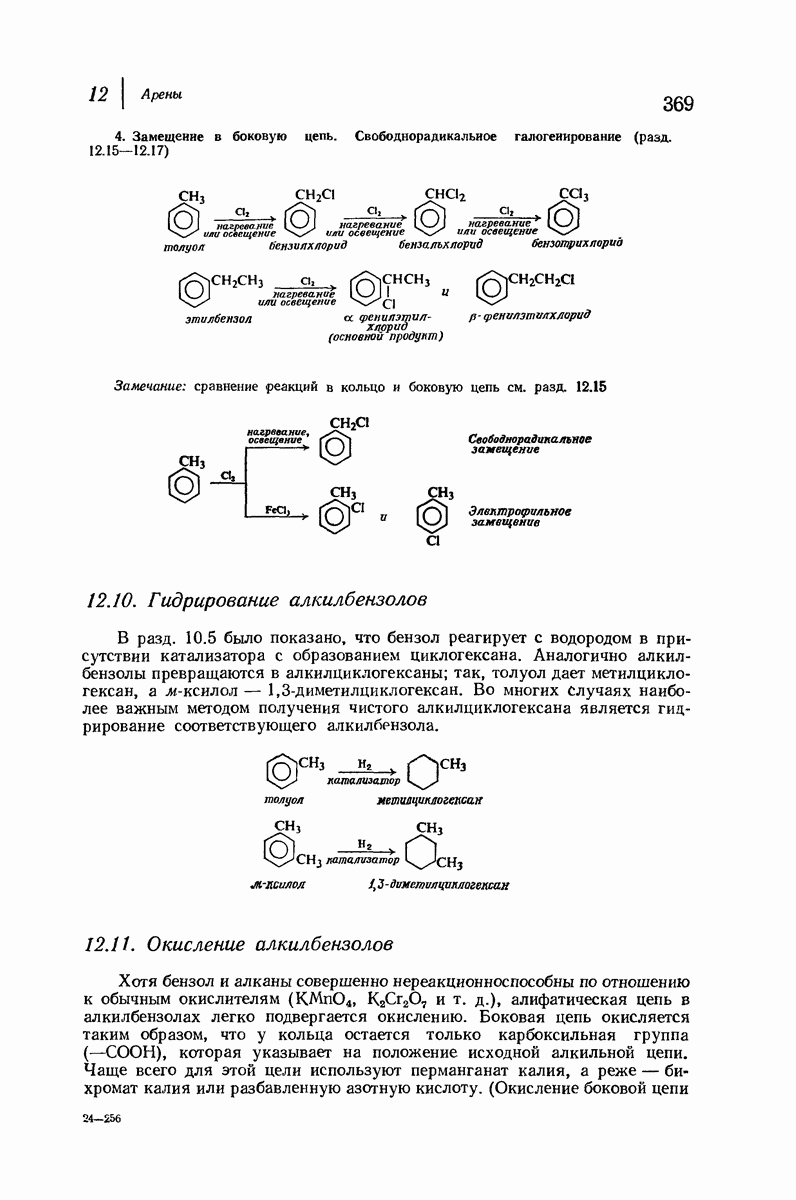
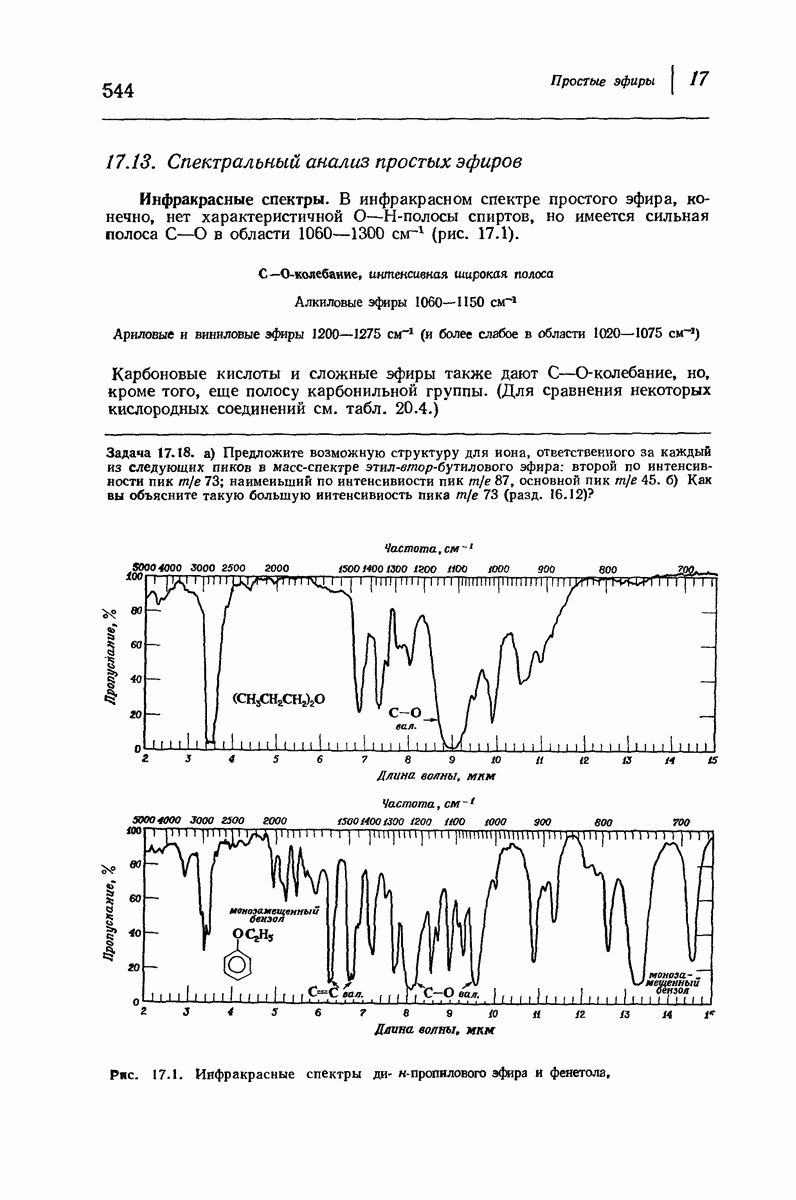
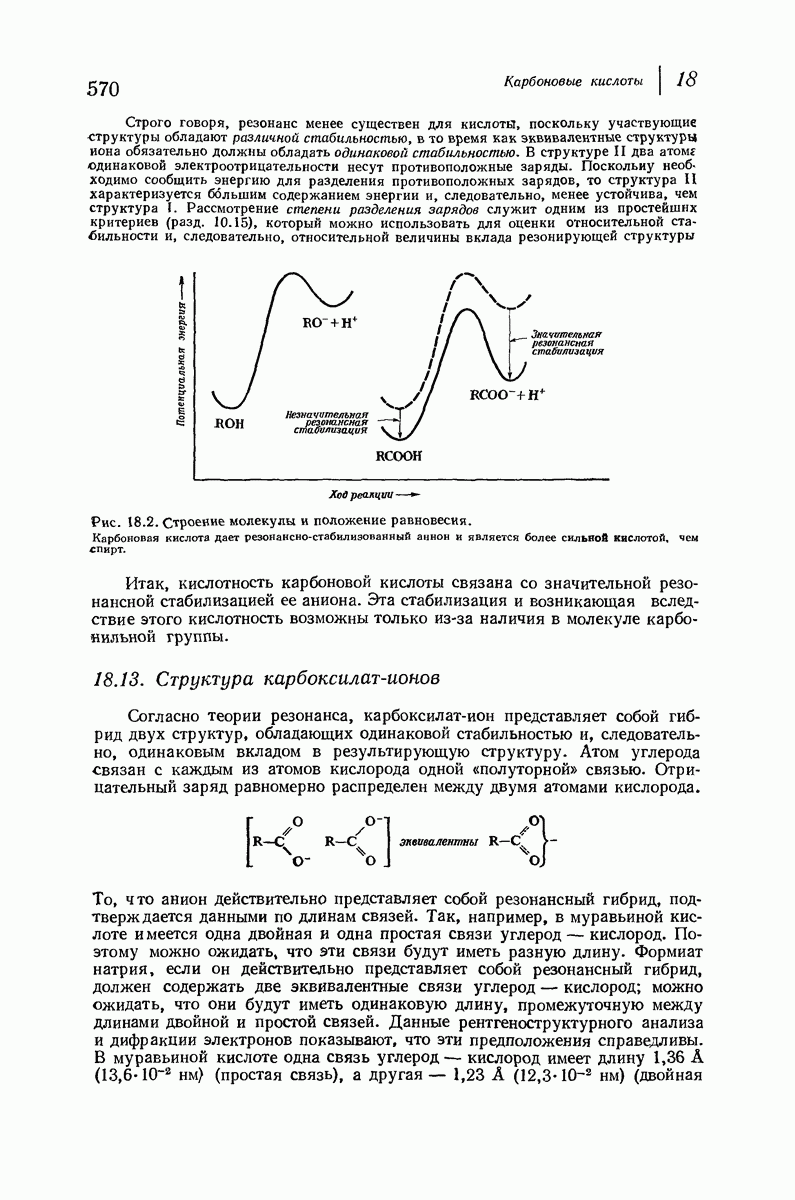
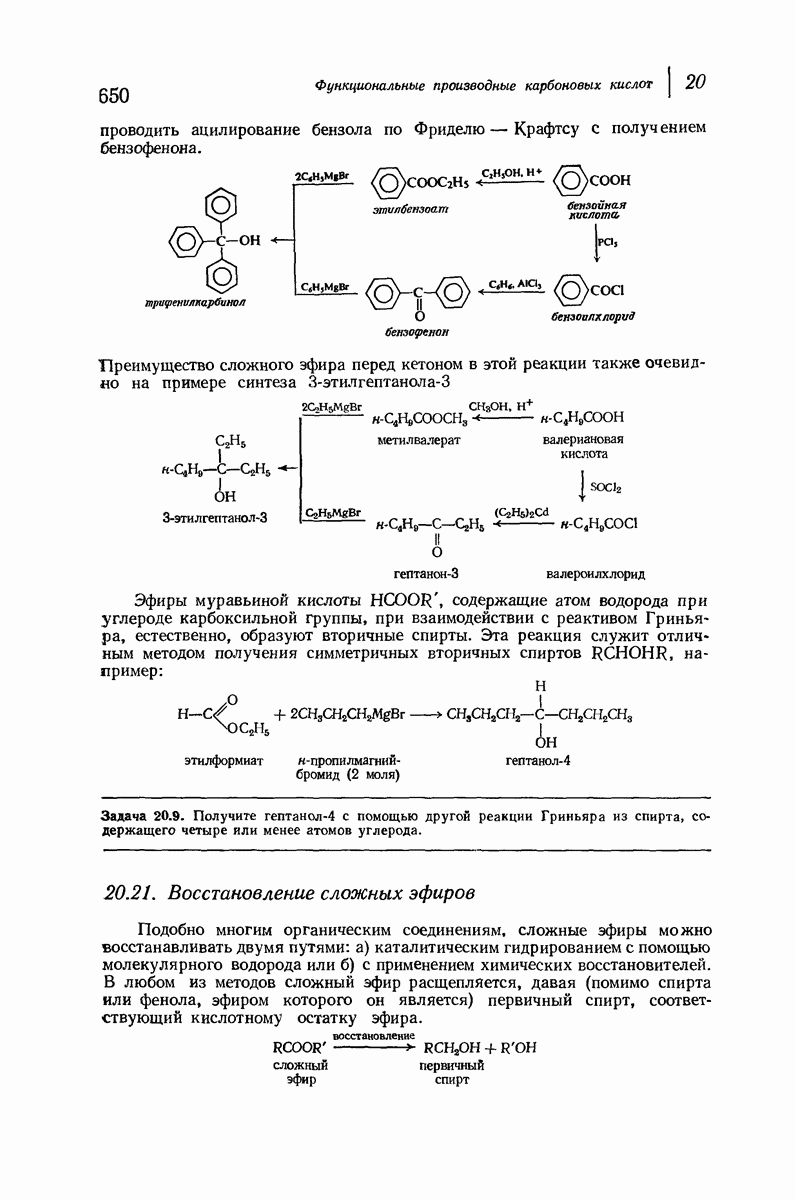
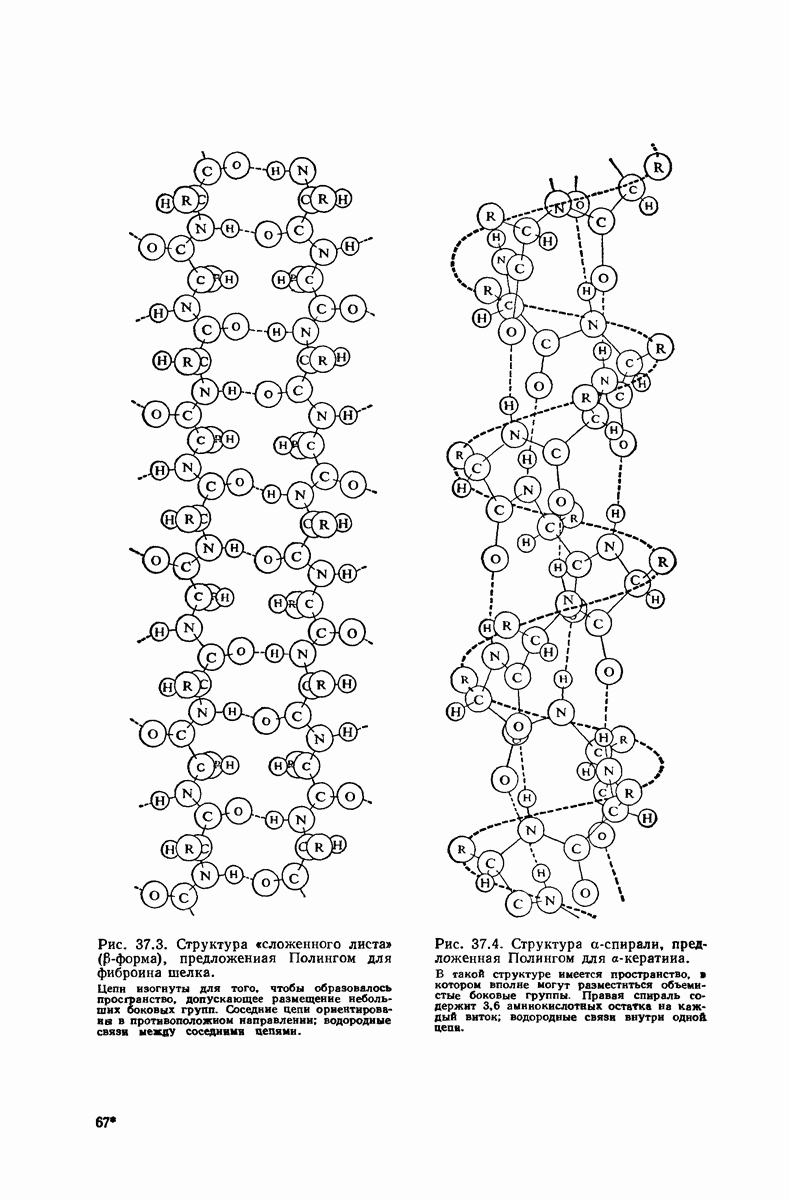
Подобная структура, называемая  -формой, была предложена для фиброина шелка, в котором период идентичности составляет 7,0 А
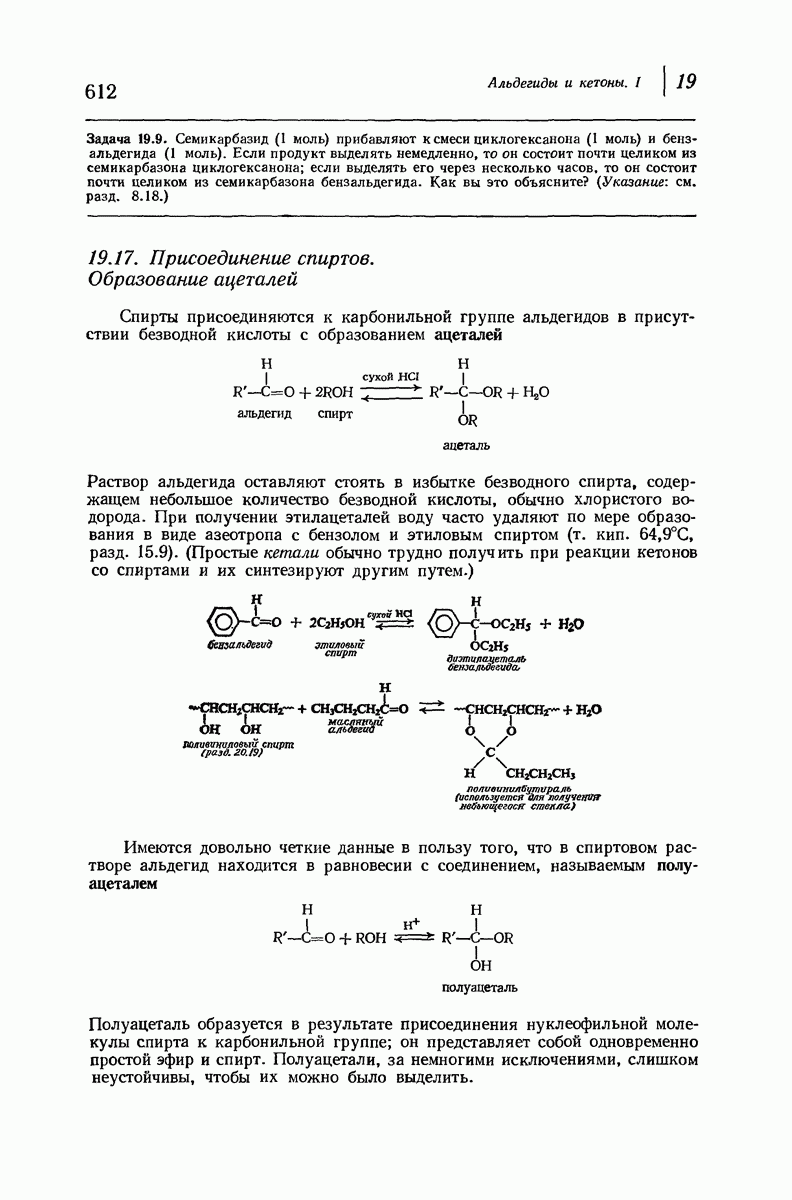
-формой, была предложена для фиброина шелка, в котором период идентичности составляет 7,0 А  и который по конфигурации ближе всего к полностью вытянутой плоской структуре листа. Существенно, что хотя в фиброине шелка имеется около 15 типов аминокислот, 46% звеньев составляет глицин, не имеющий боковых групп, а еще 38% — аланин и серин с небольшими боковыми группами —
и который по конфигурации ближе всего к полностью вытянутой плоской структуре листа. Существенно, что хотя в фиброине шелка имеется около 15 типов аминокислот, 46% звеньев составляет глицин, не имеющий боковых групп, а еще 38% — аланин и серин с небольшими боковыми группами —  и
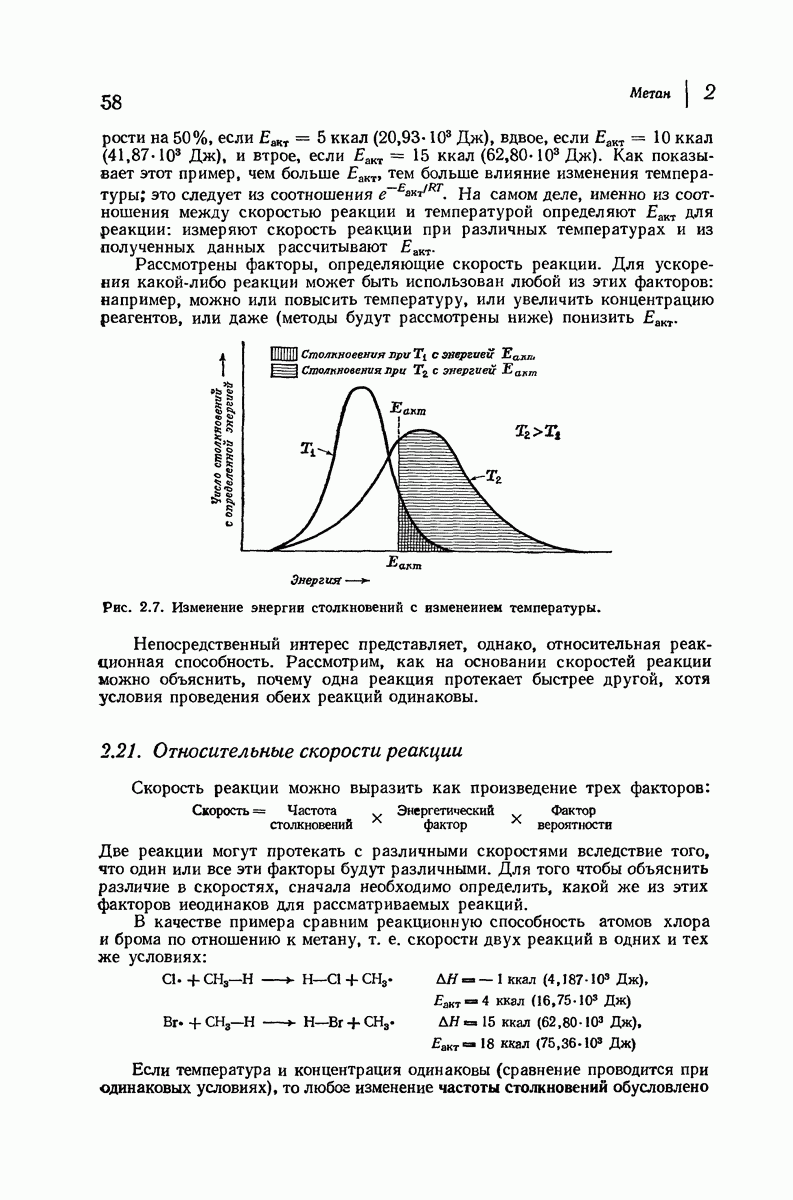
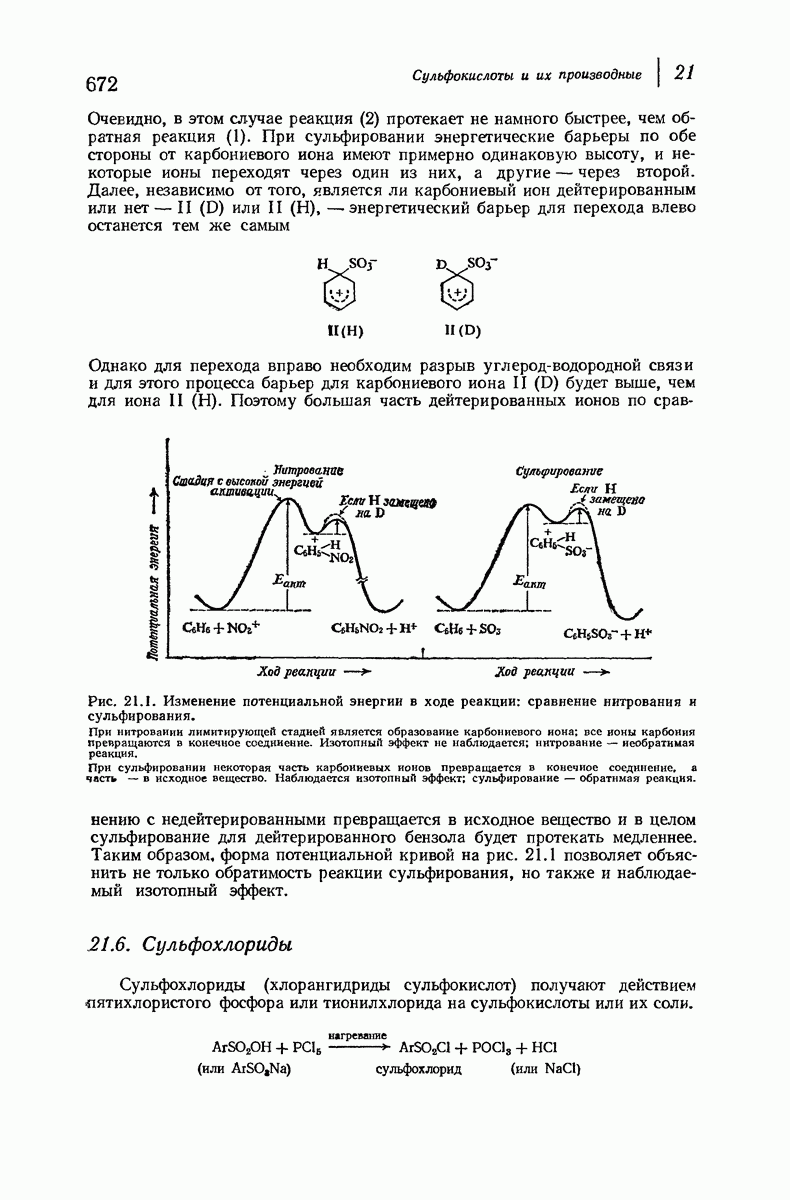
и 
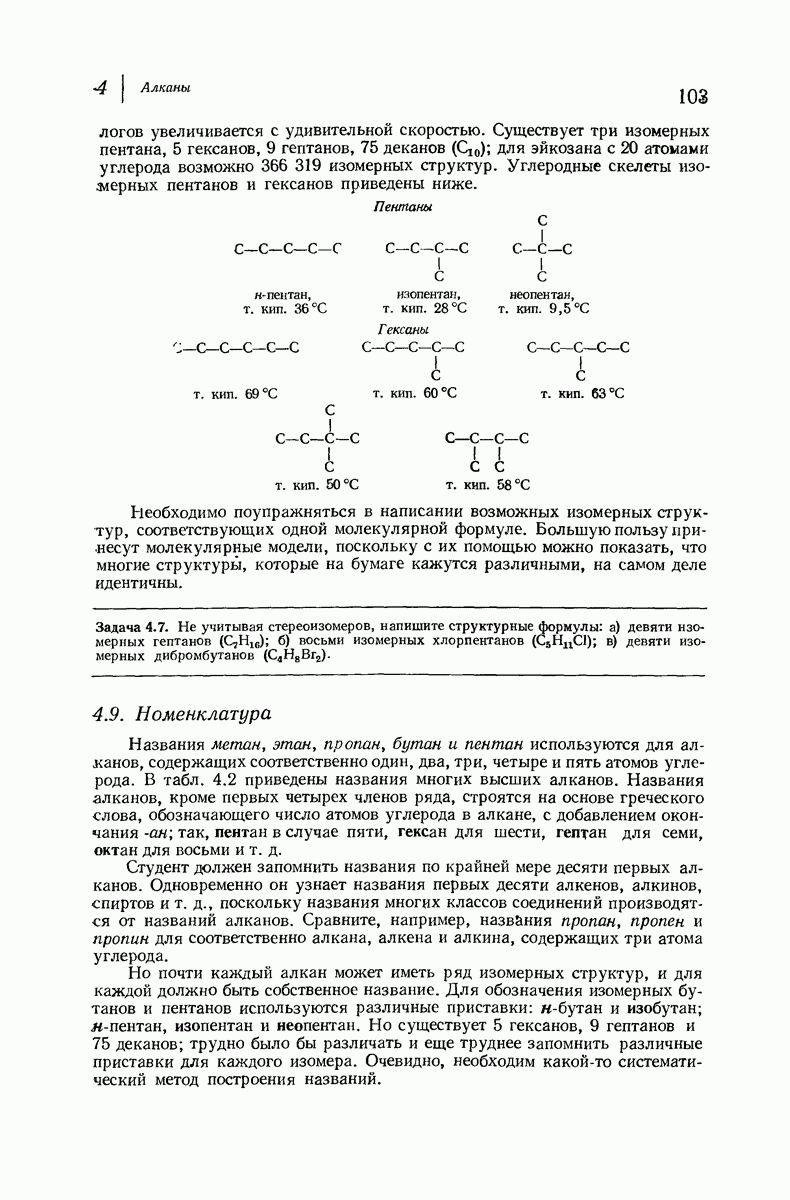
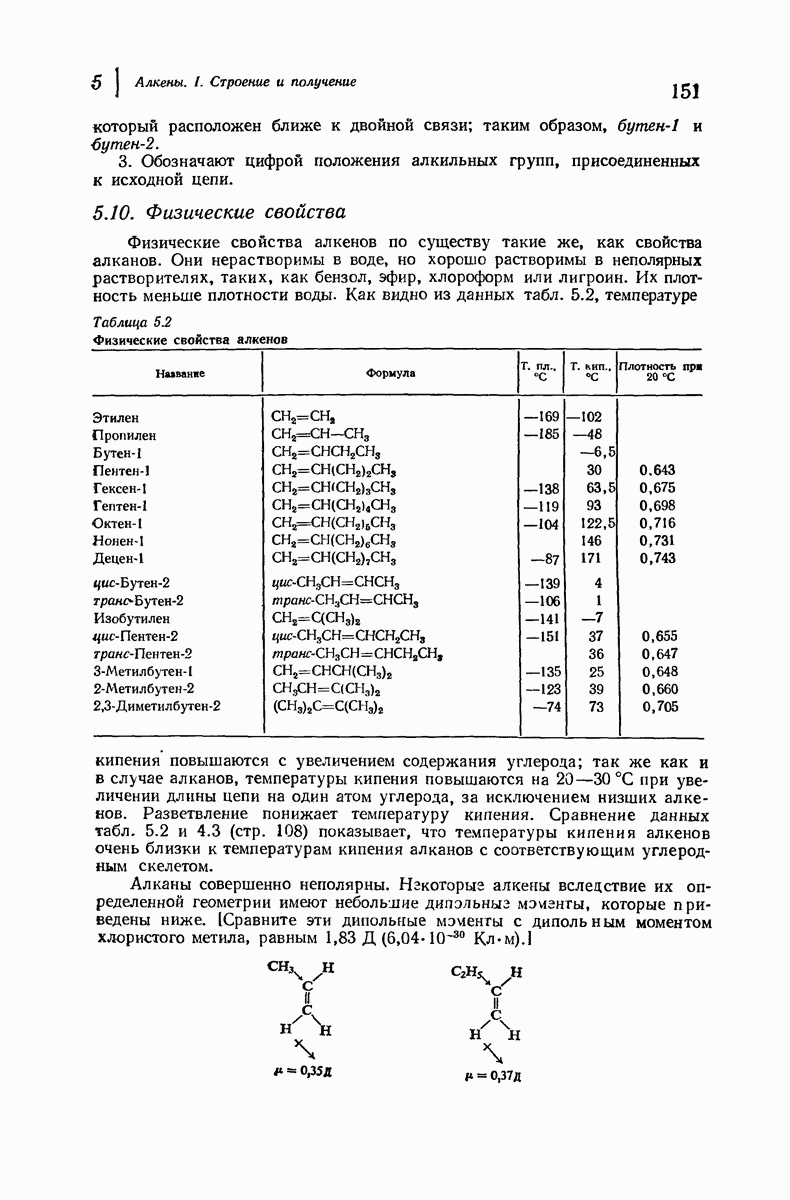
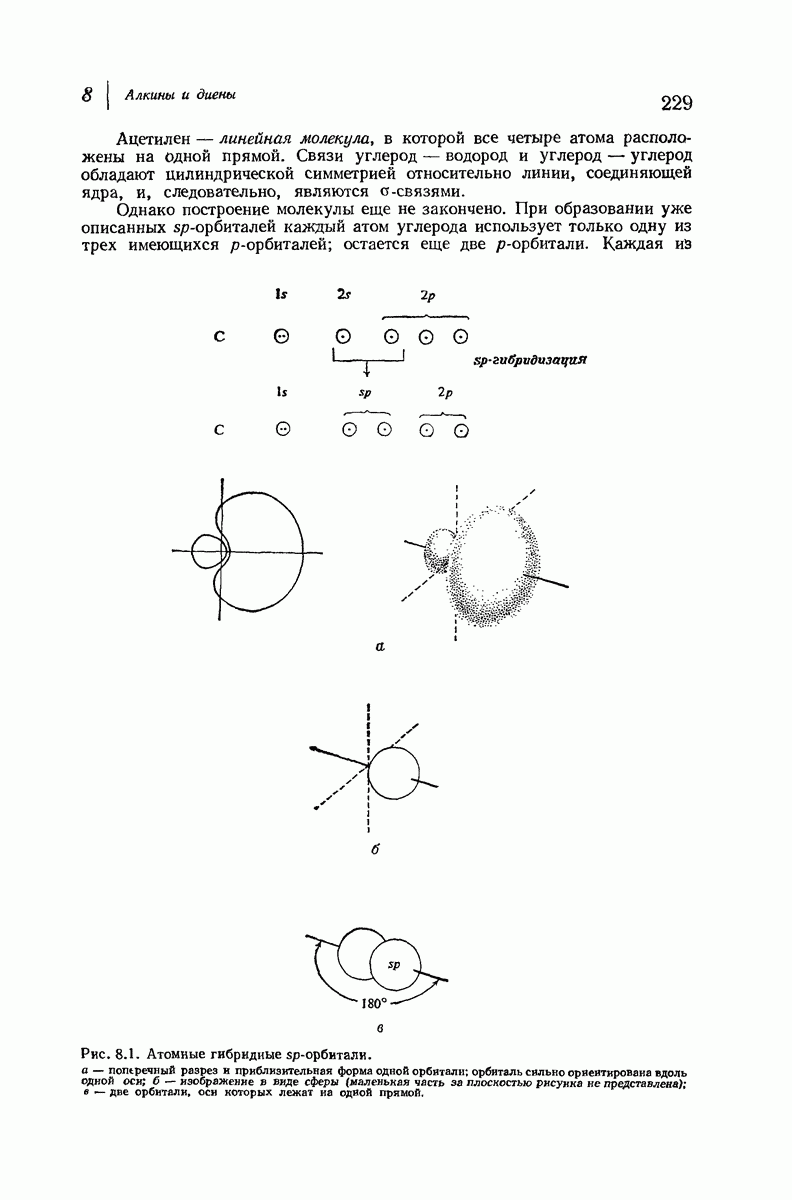
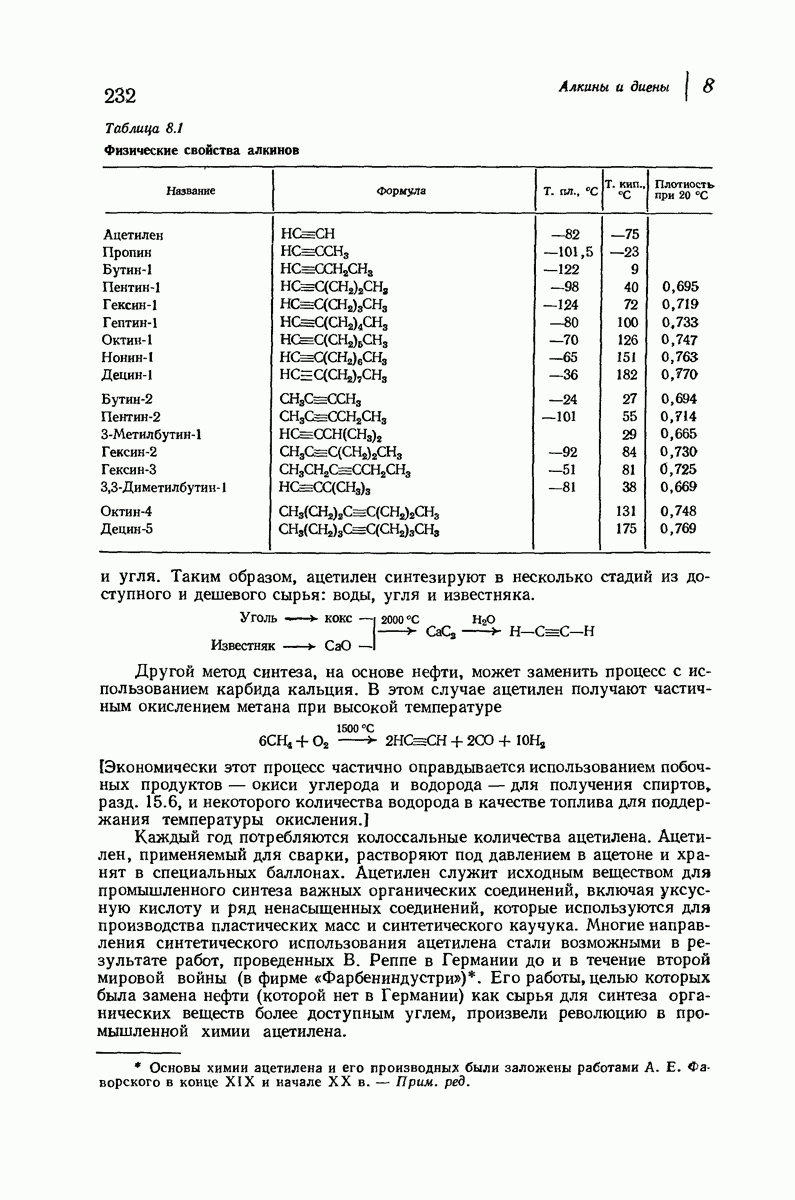
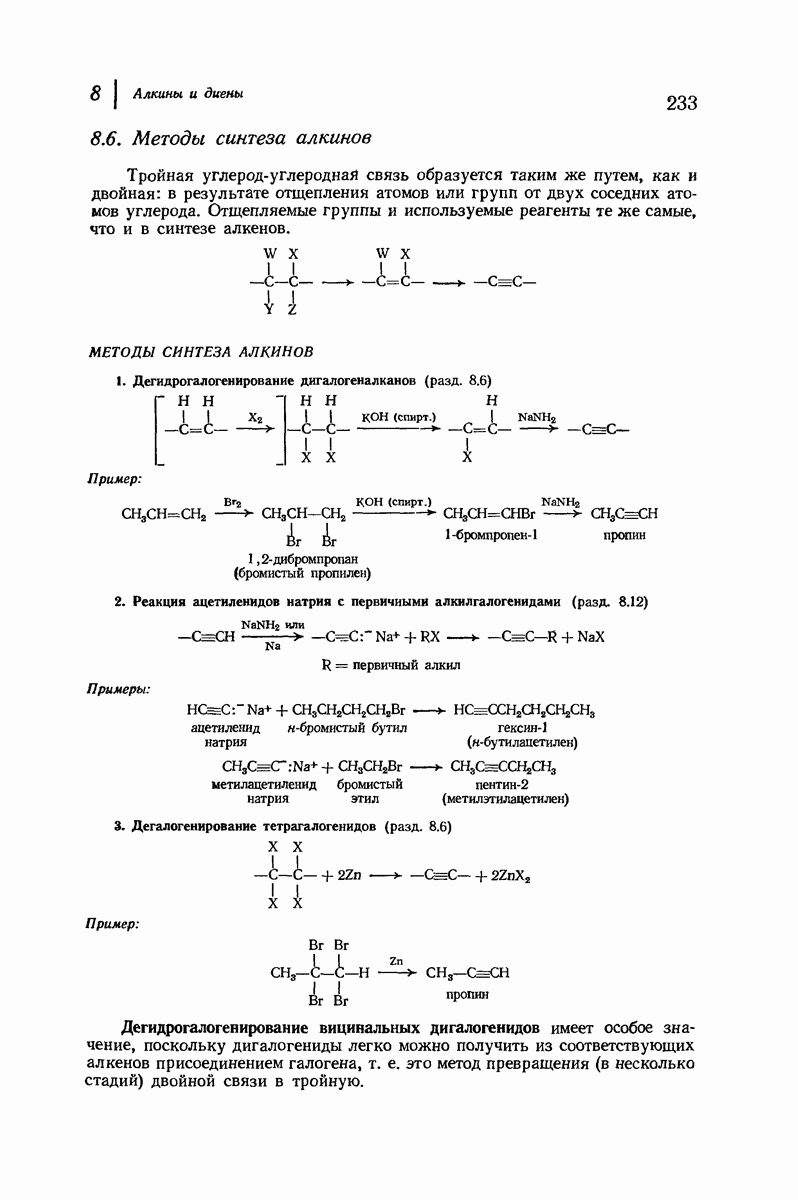
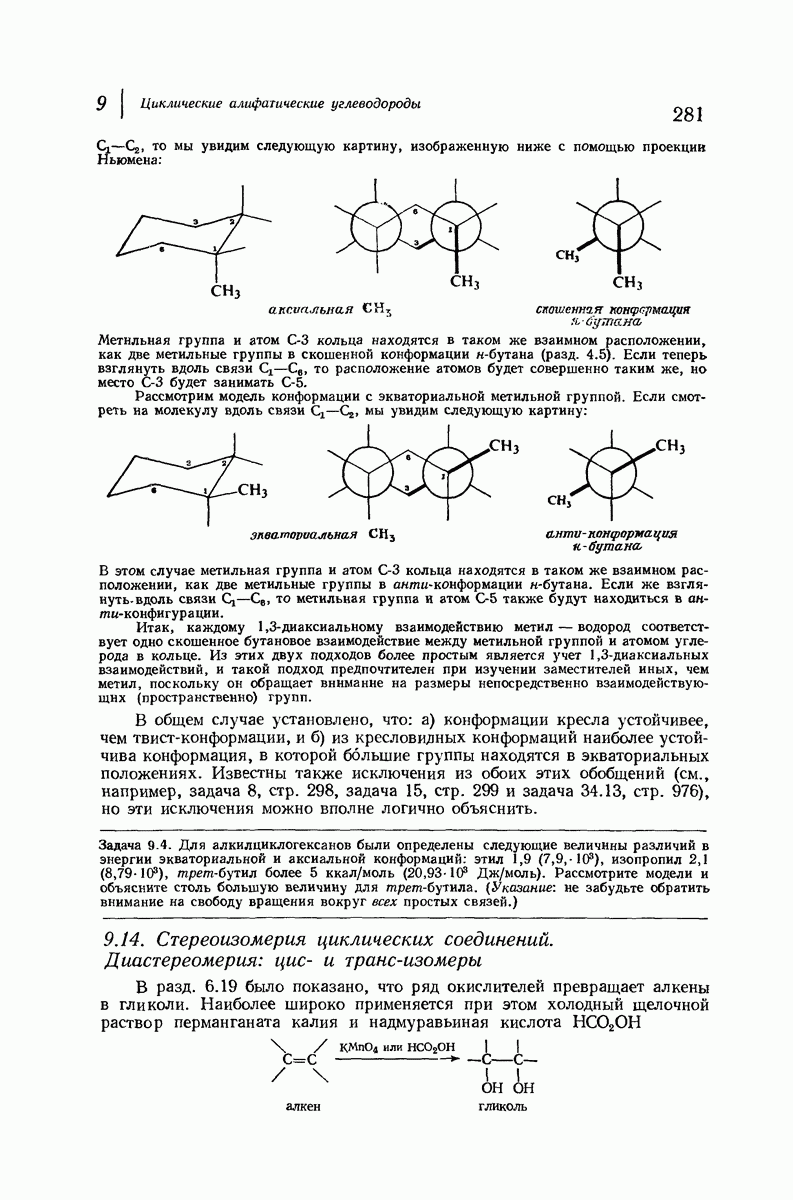
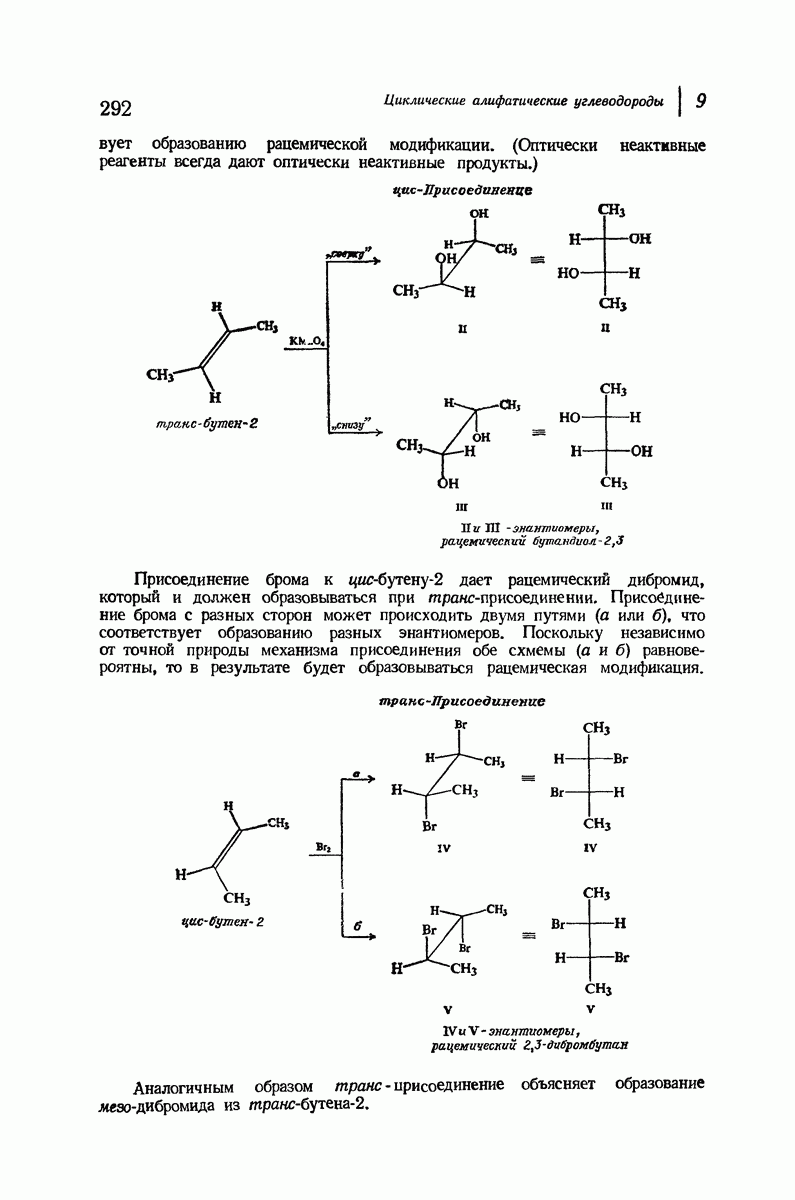
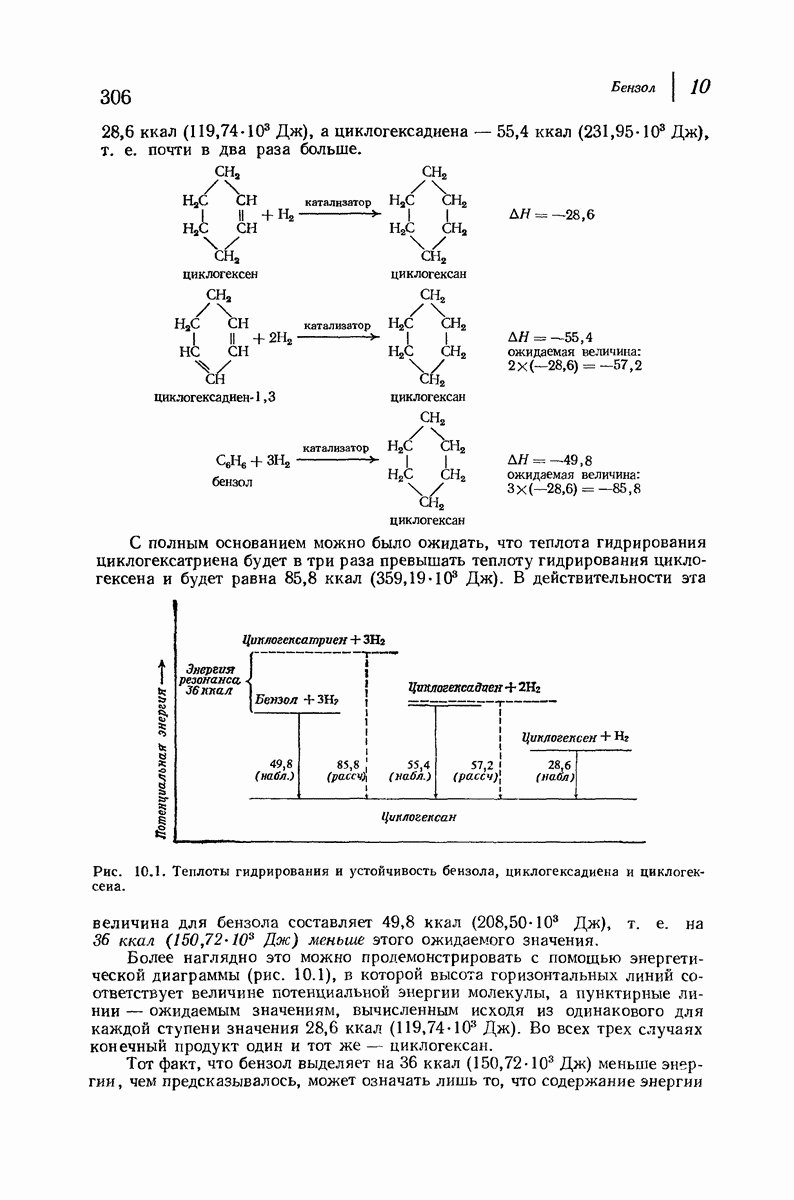
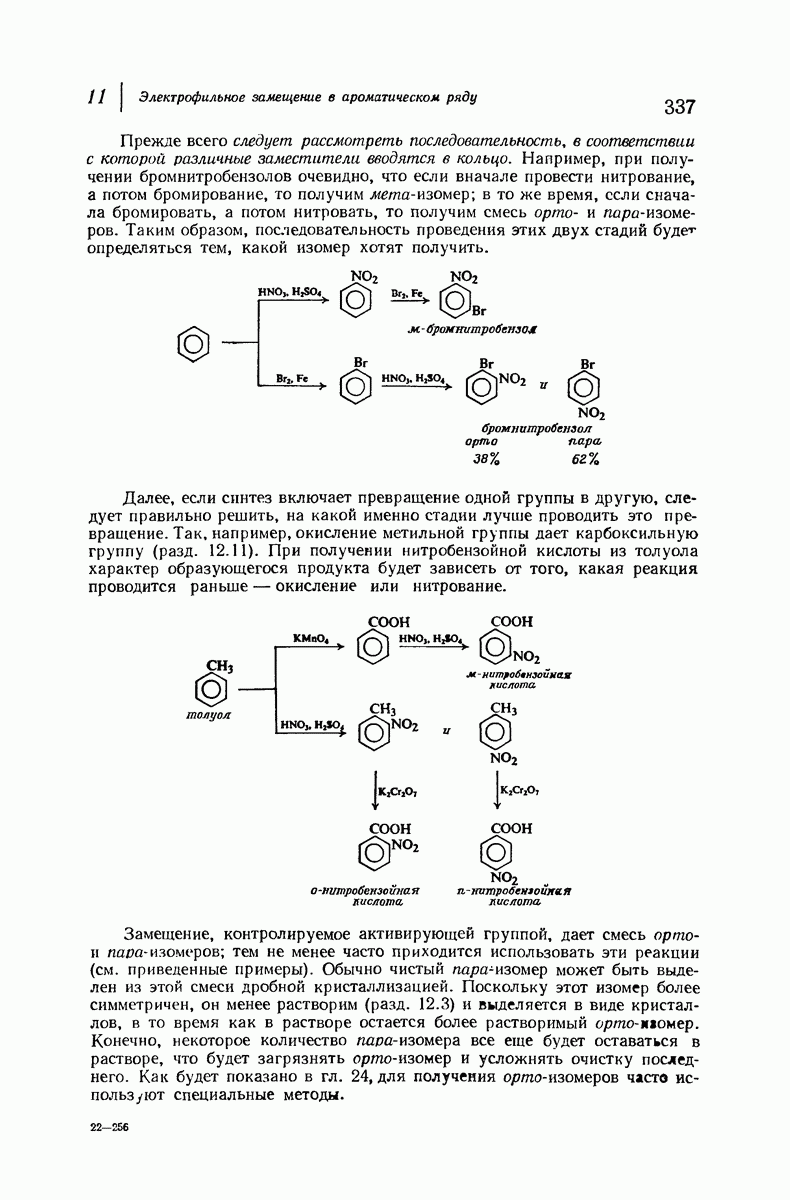
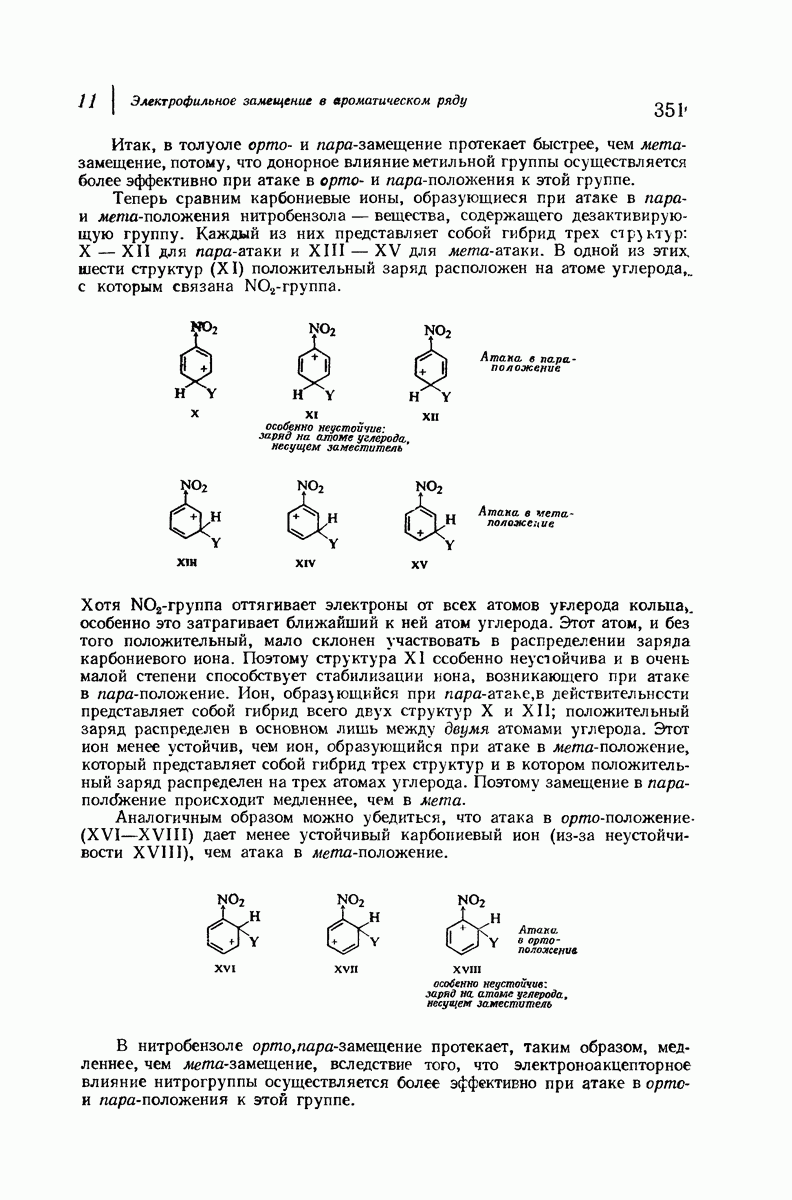
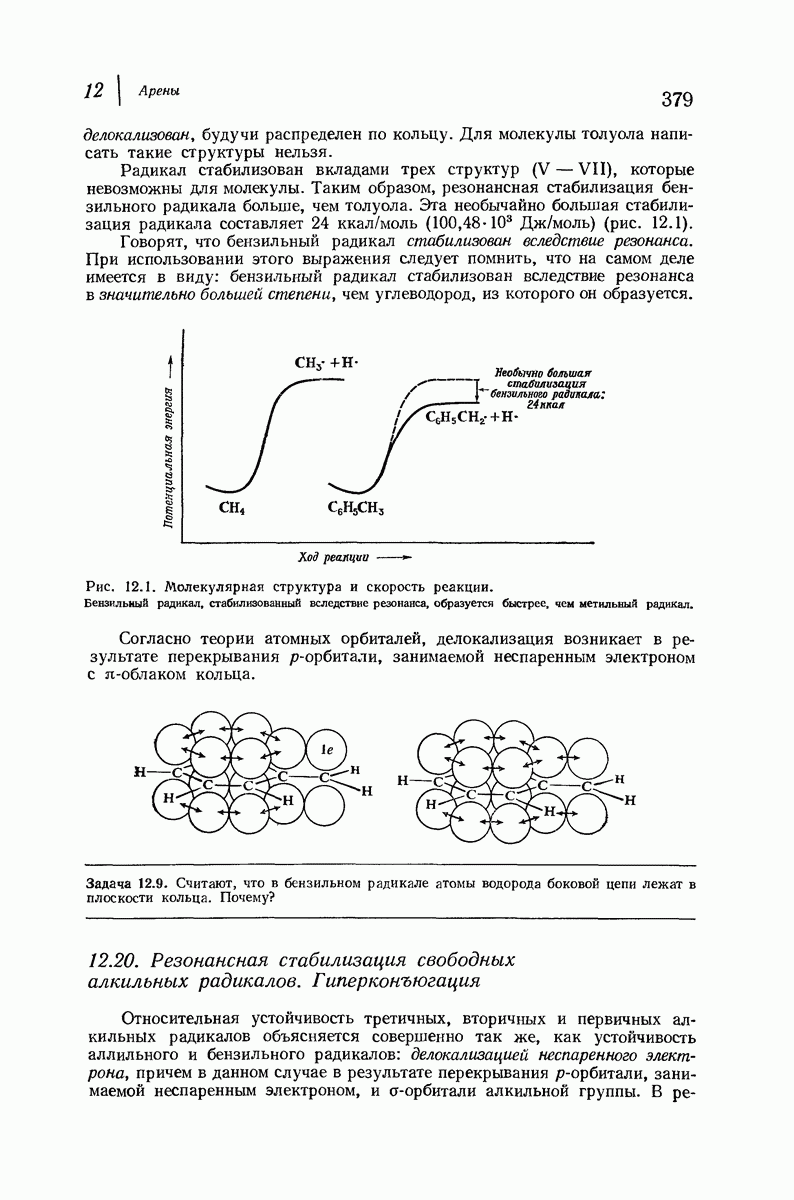
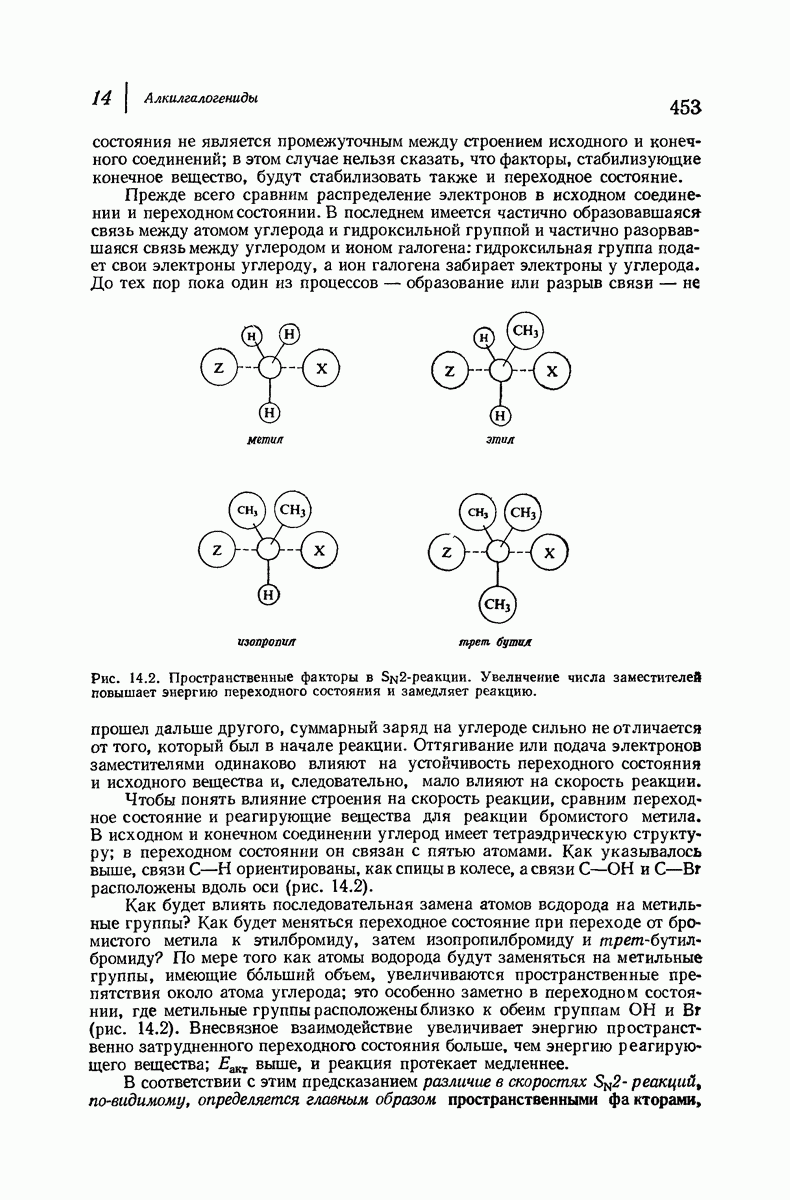
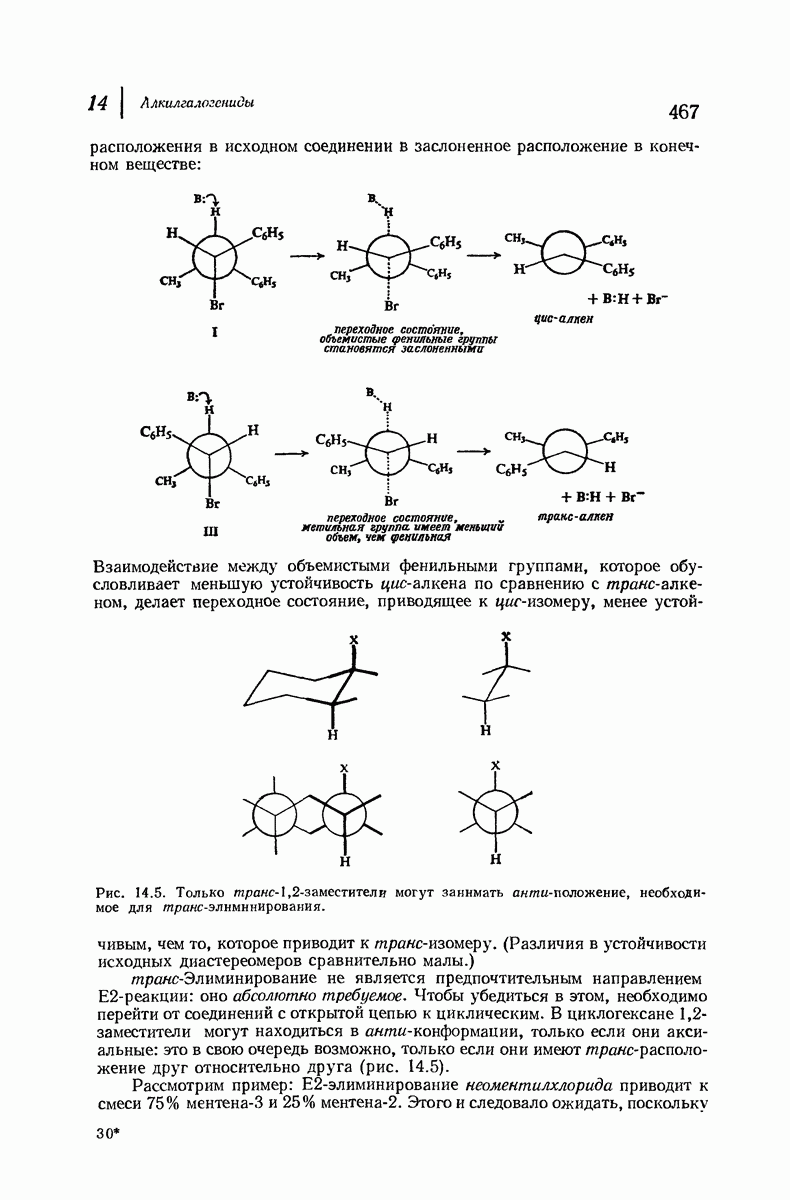
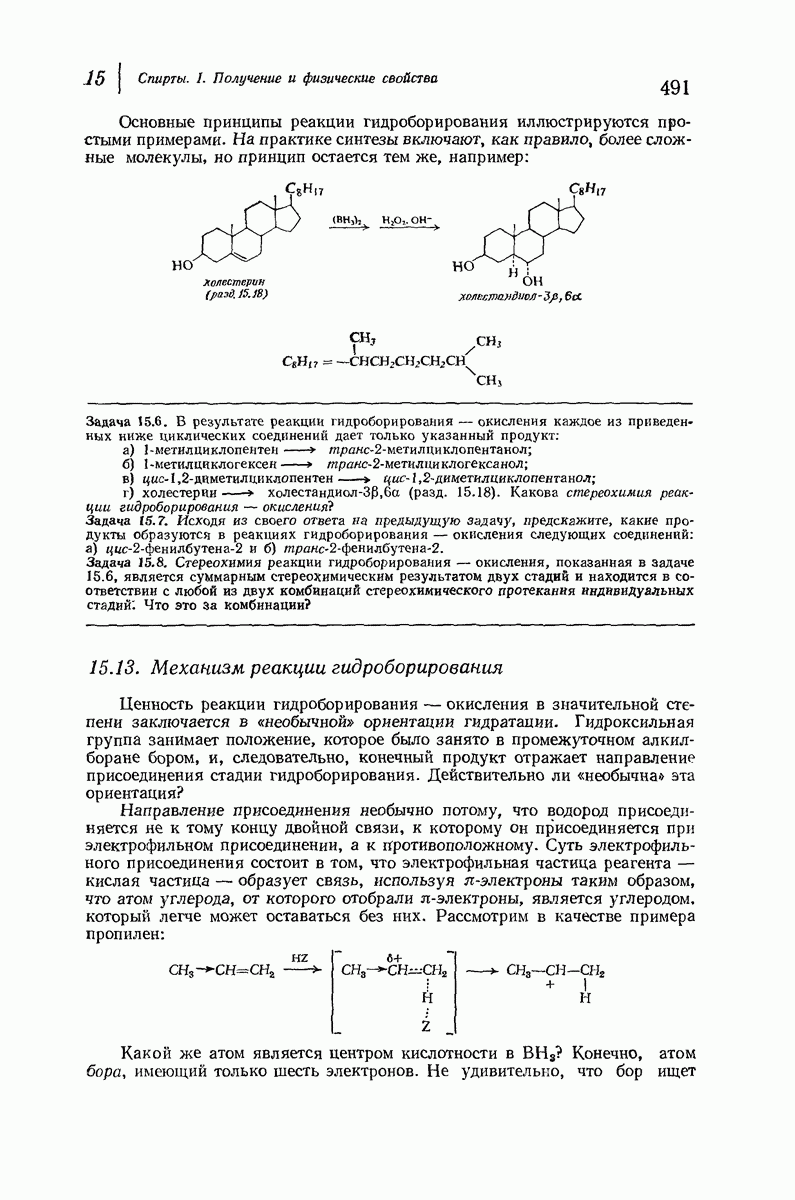
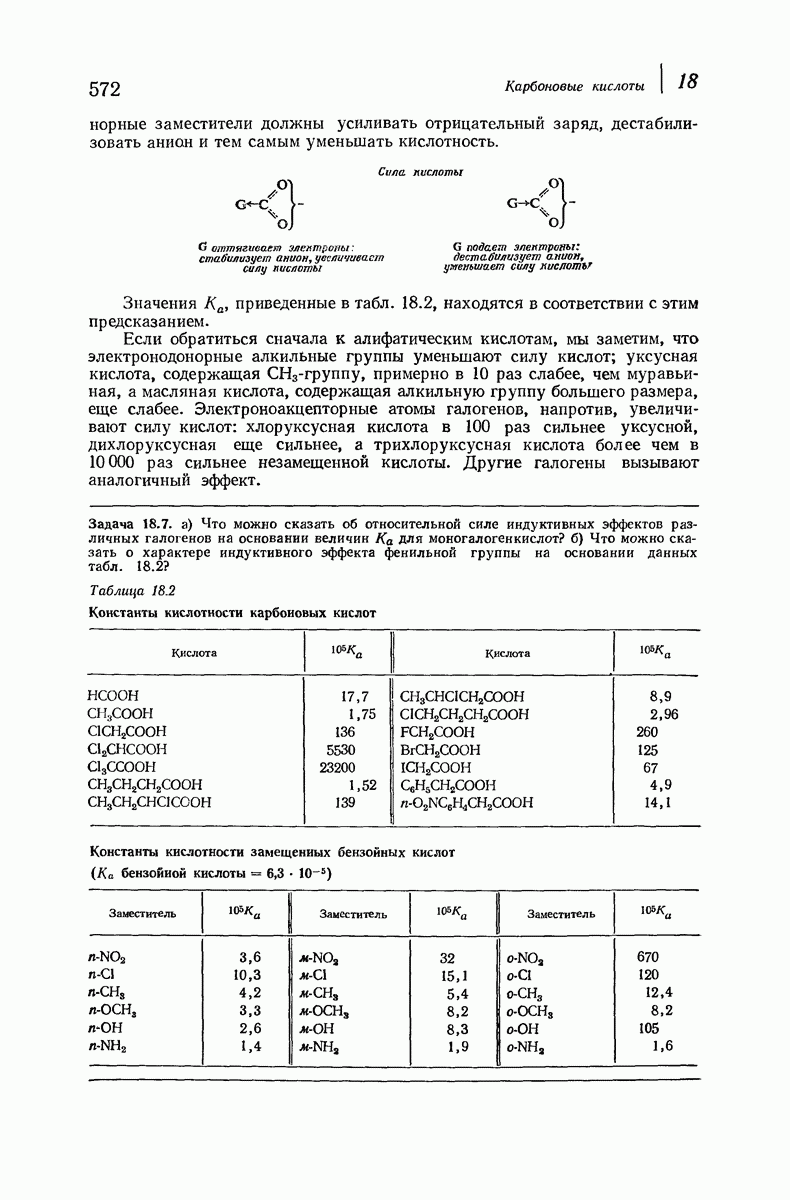
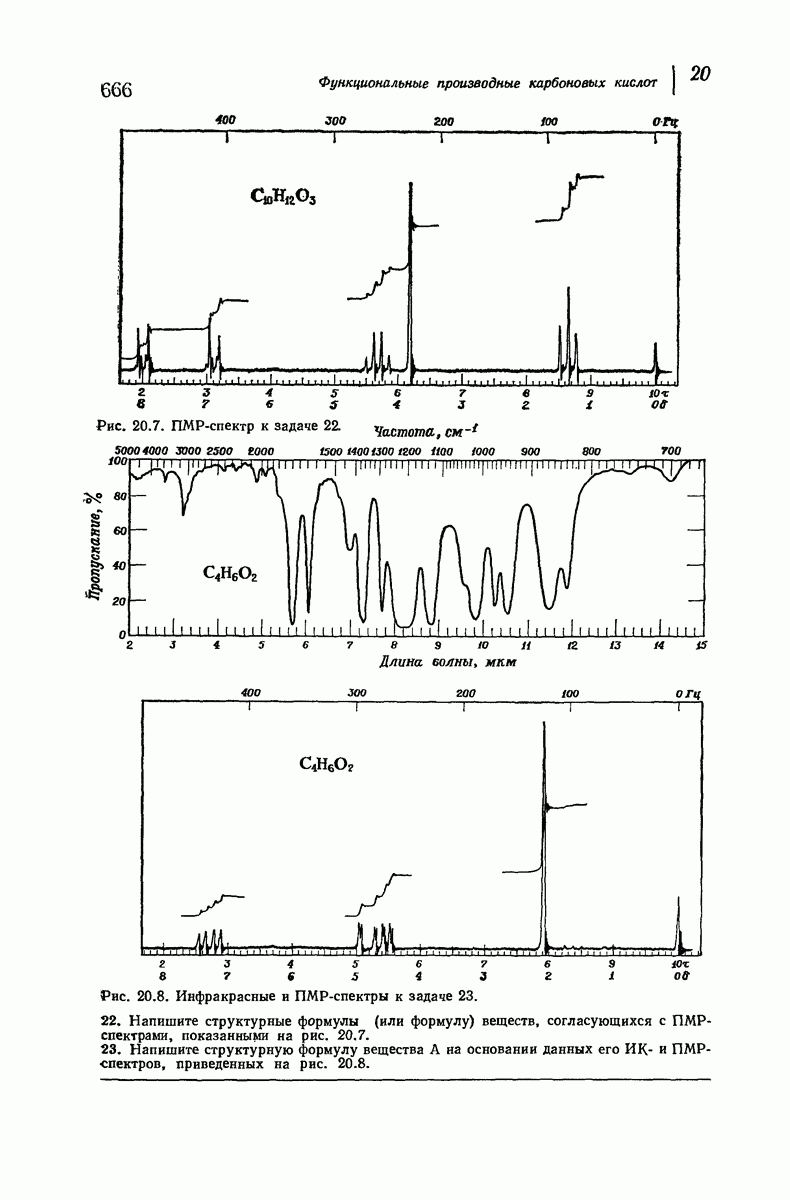
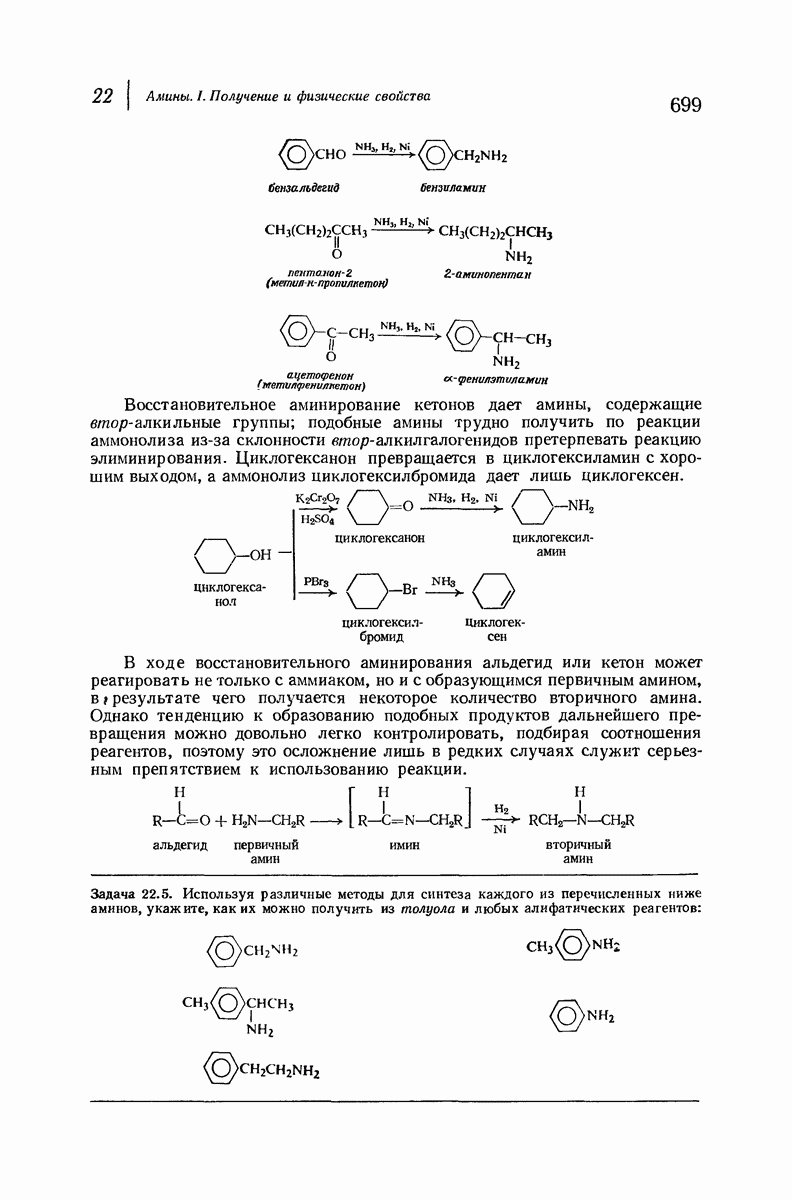
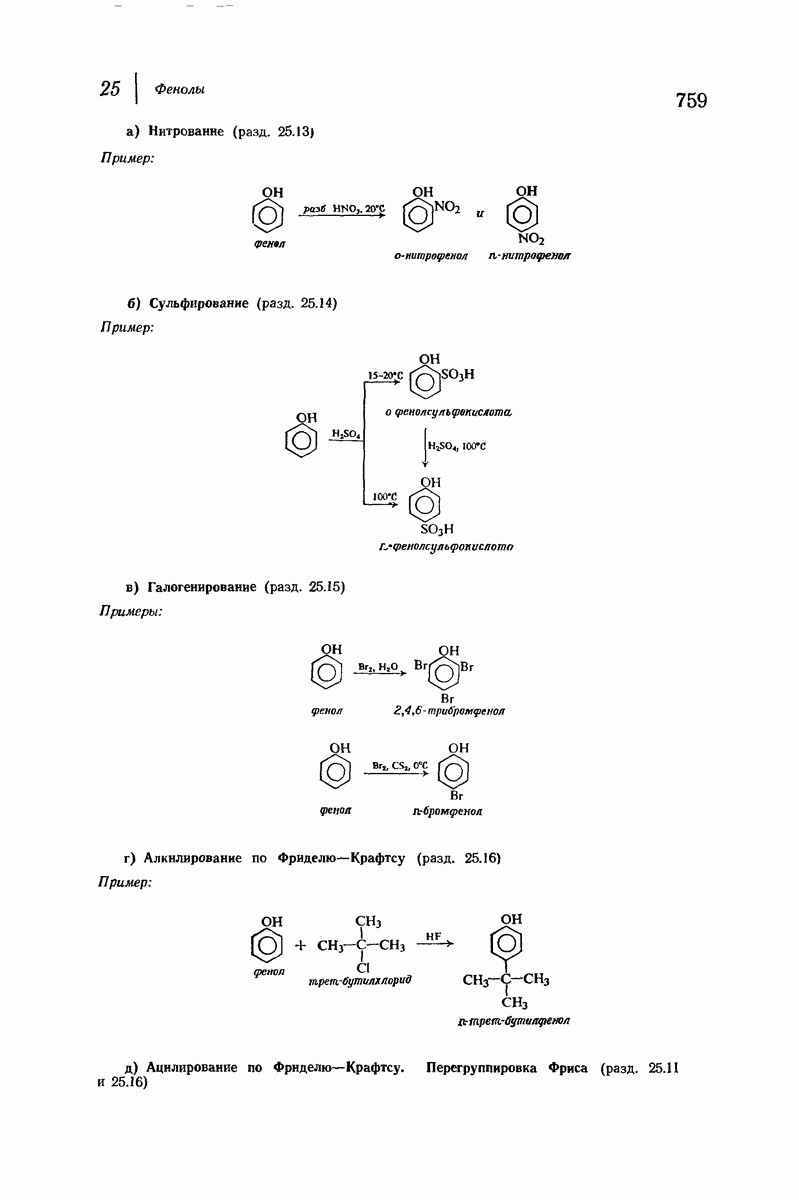
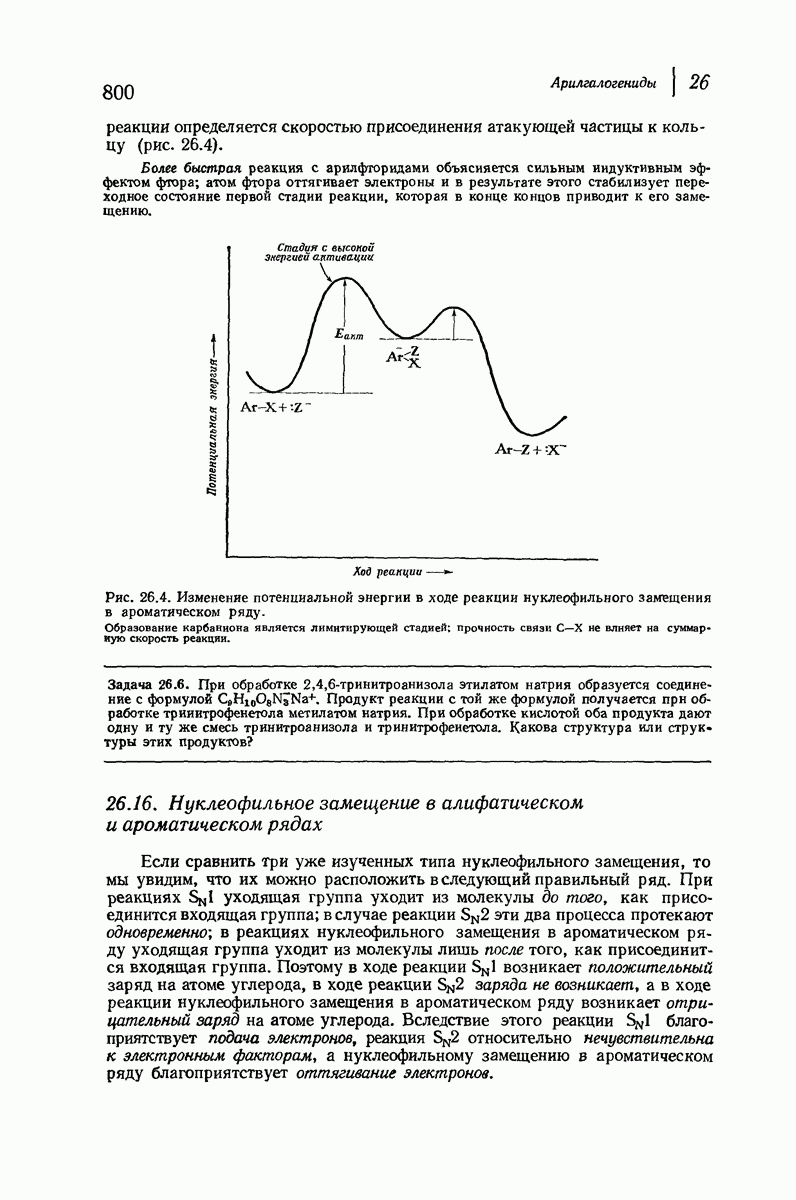
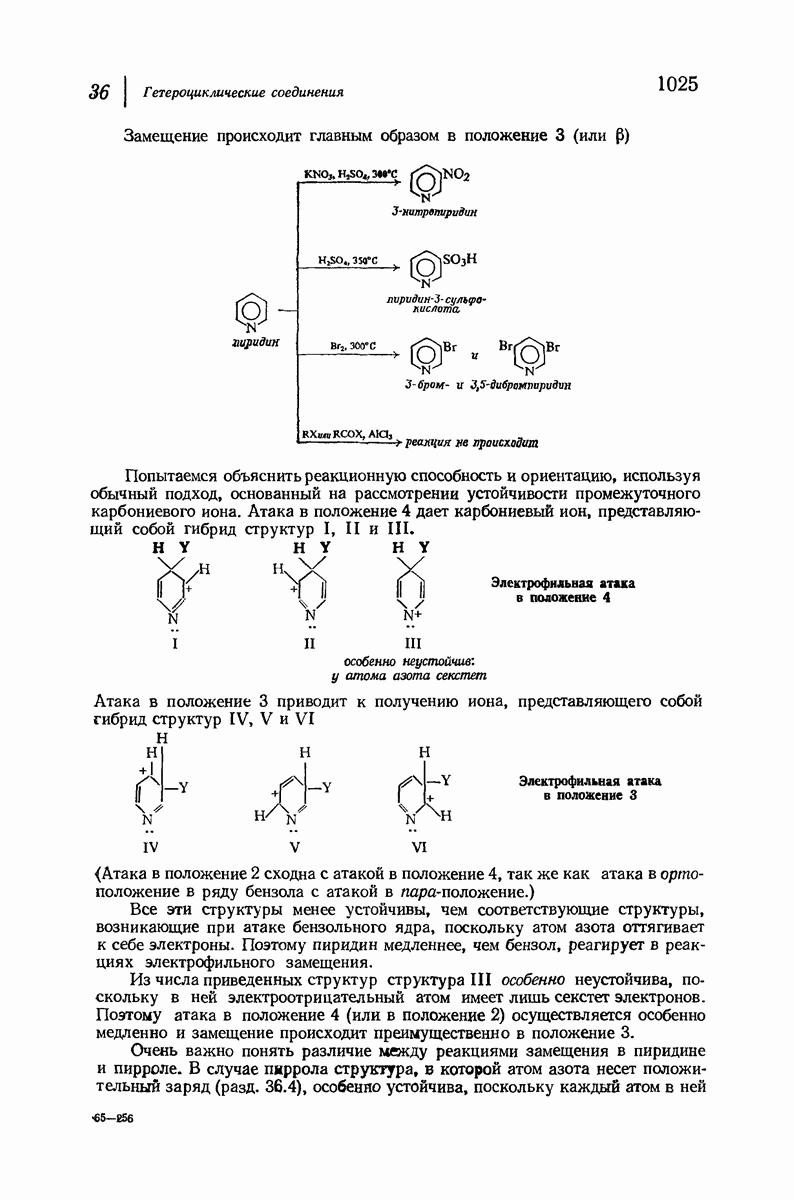
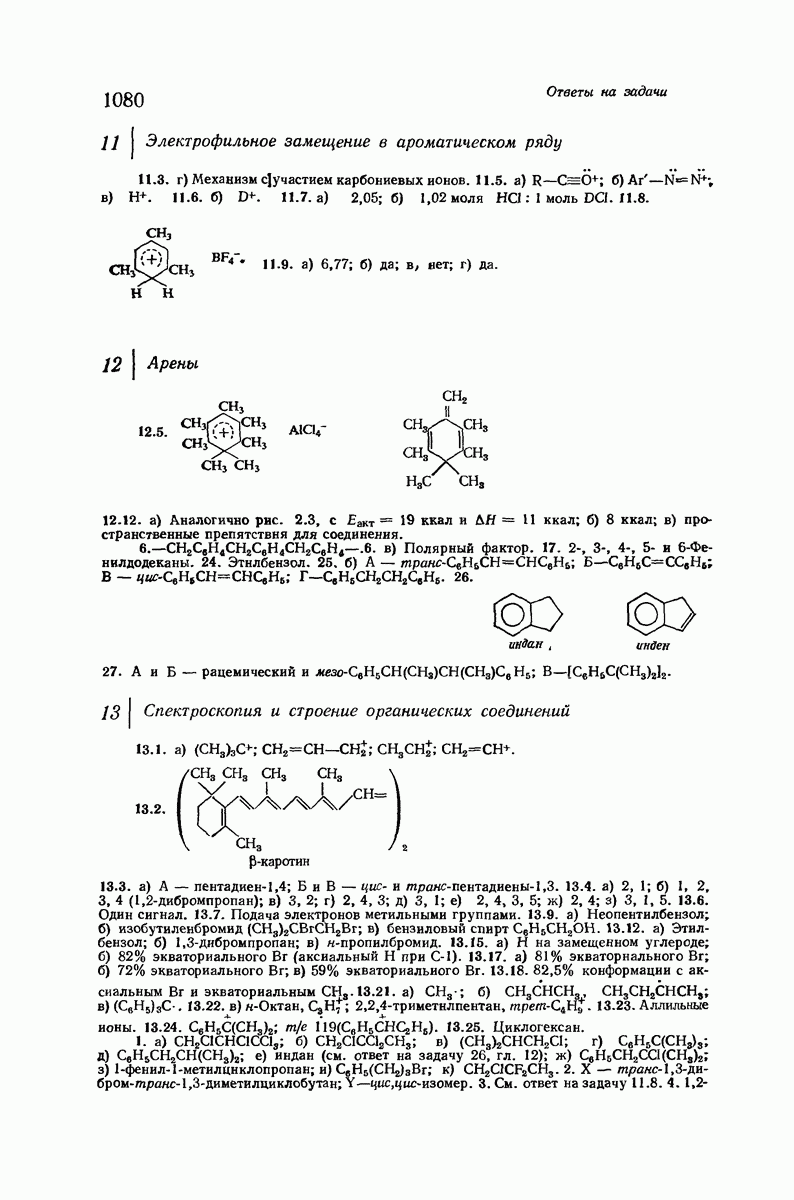
Боковые группы большого размера лучше всего могут быть размещены в структуре совсем иного рода. Каждая цепь закручена так, что образуется спираль (как винтовая лестница). Между различными участками одной и той же цепи возникают водородные связи, фиксирующие структуру спирали. Для  -кератина (нерастянутая шерсть, волосы, рога, ногти) Полинг предложил спираль, в которой на один виток (шаг спирали) приходится 3,6 аминокислотных остатка (рис. 37.4). Согласно моделям, подобная спираль, содержащая 3,6 аминокислотных остатка на виток, образует пространство, достаточное для размещения боковых цепочек, и позволяет образоваться
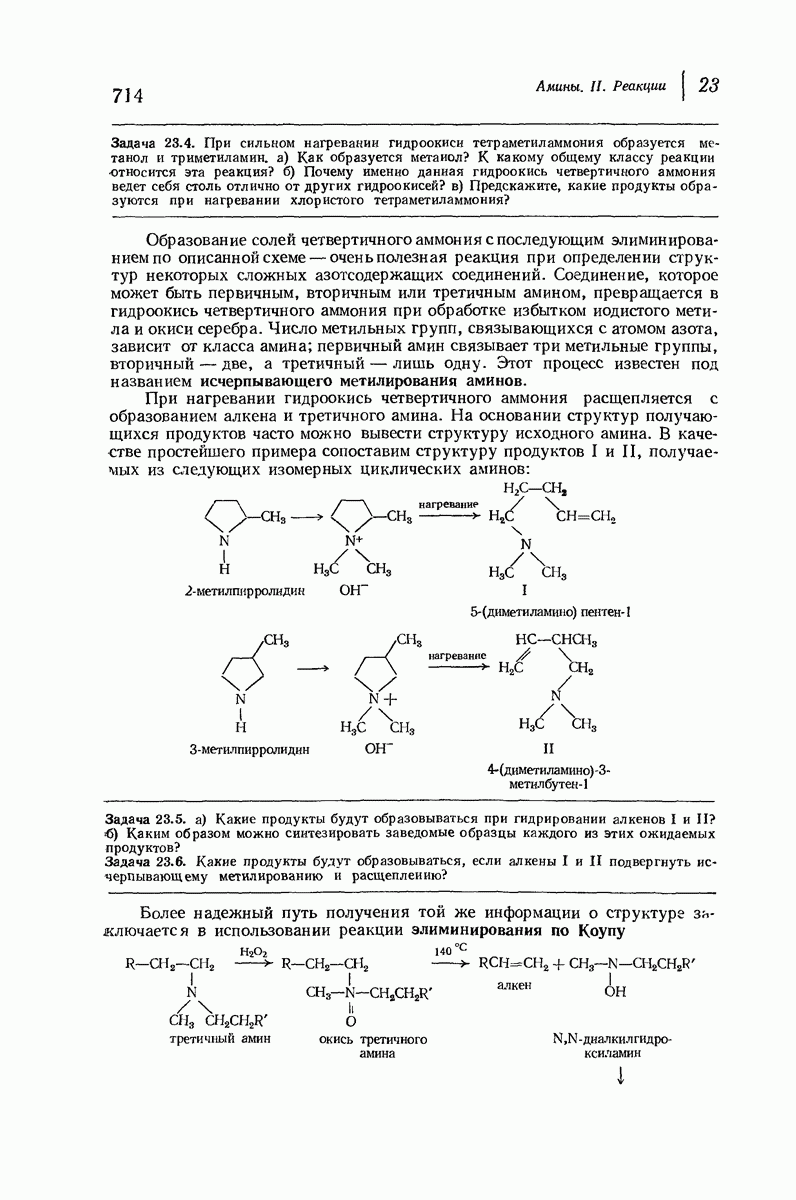
-кератина (нерастянутая шерсть, волосы, рога, ногти) Полинг предложил спираль, в которой на один виток (шаг спирали) приходится 3,6 аминокислотных остатка (рис. 37.4). Согласно моделям, подобная спираль, содержащая 3,6 аминокислотных остатка на виток, образует пространство, достаточное для размещения боковых цепочек, и позволяет образоваться

(кликните для просмотра скана)

всевозможным водородным связям. Она также объясняет наблюдаемую величину 1,5 А  для периодического расстояния, поскольку именно таково расстояние между аминокислотными остатками, измеренное вдоль оси спирали. Для того чтобы уложиться в такую спираль, все аминокислоты должны иметь одну и ту же конфигурацию, что и наблюдается в действительности; кроме того, поскольку они имеют
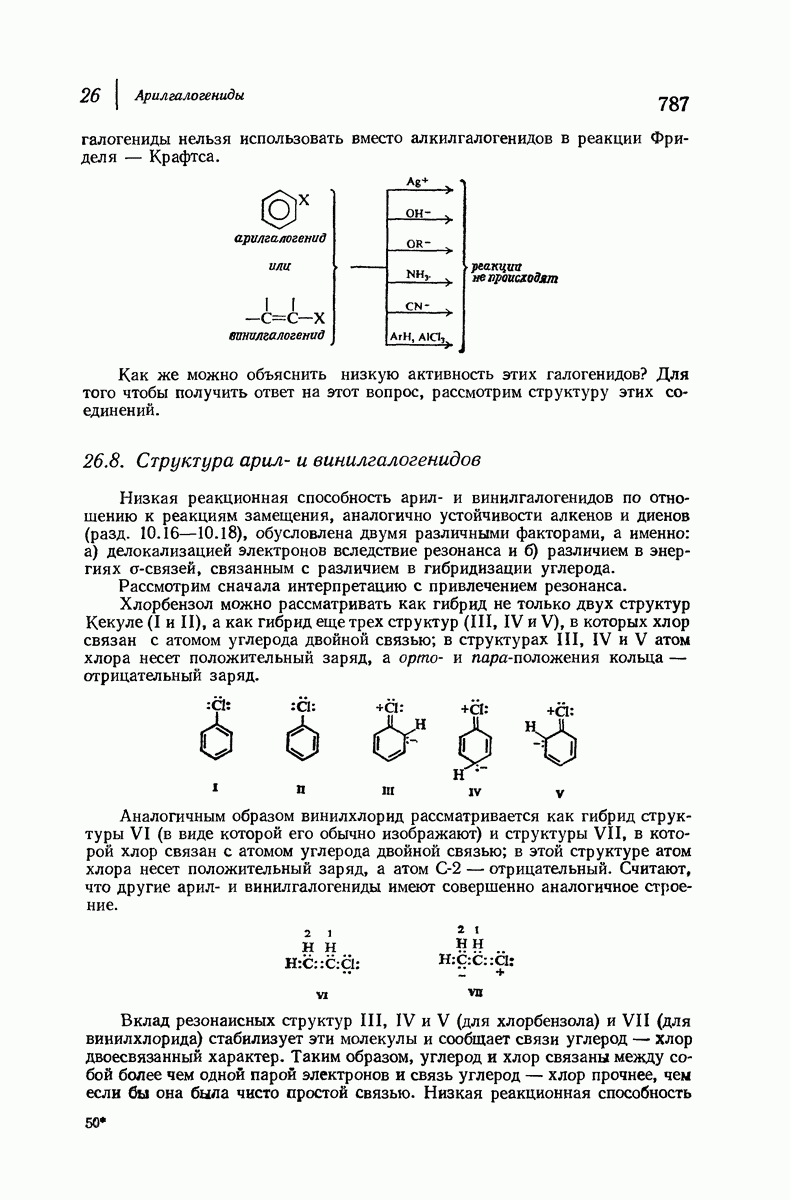
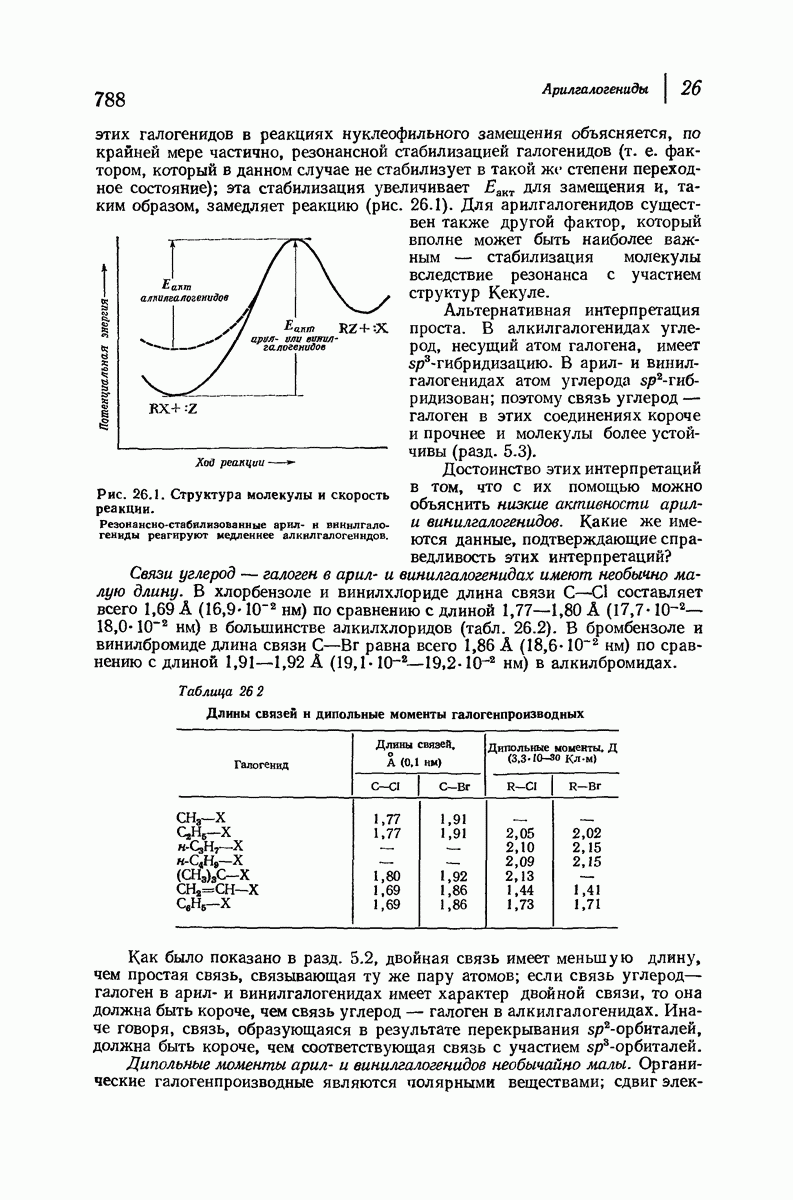
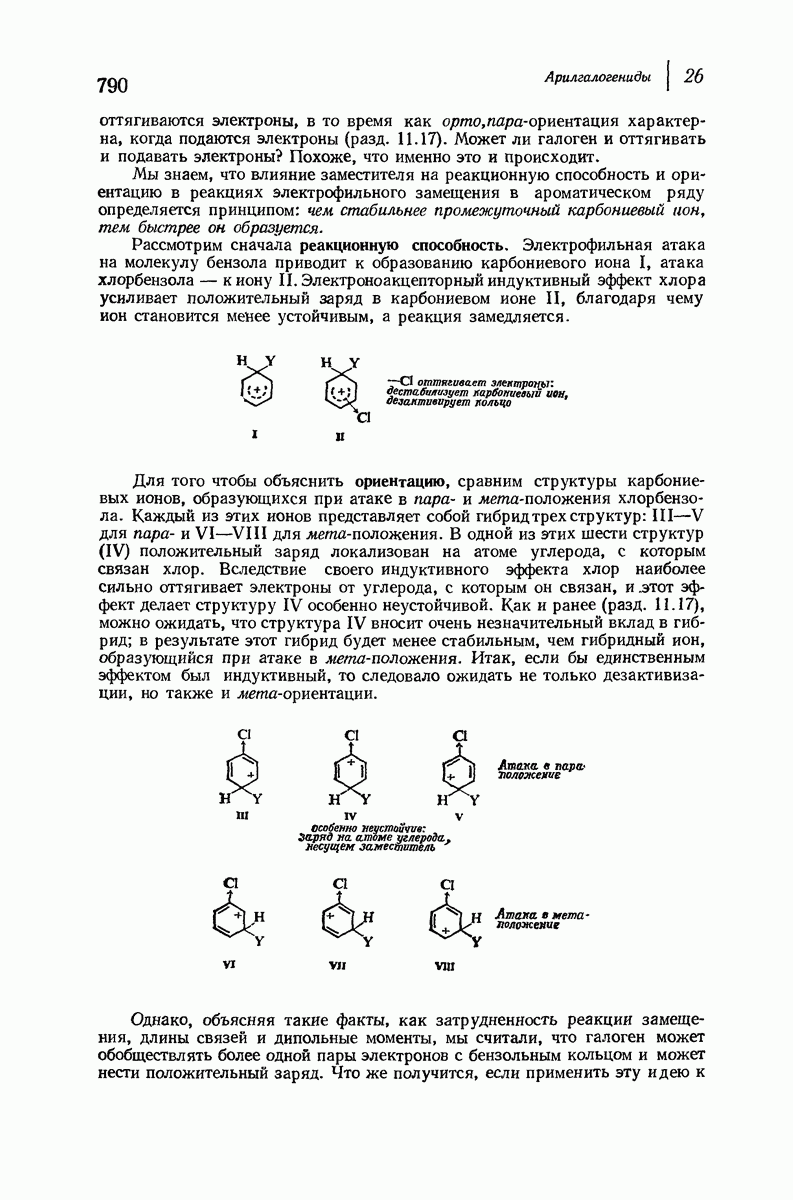
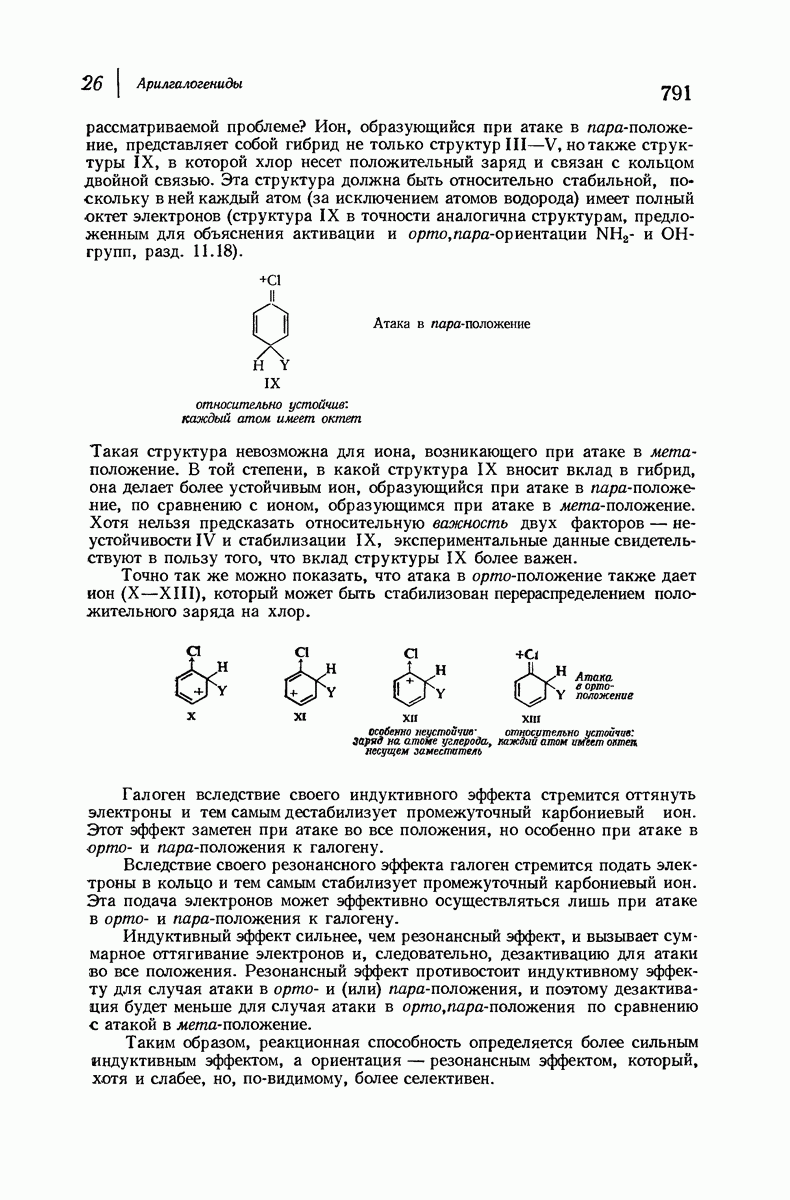
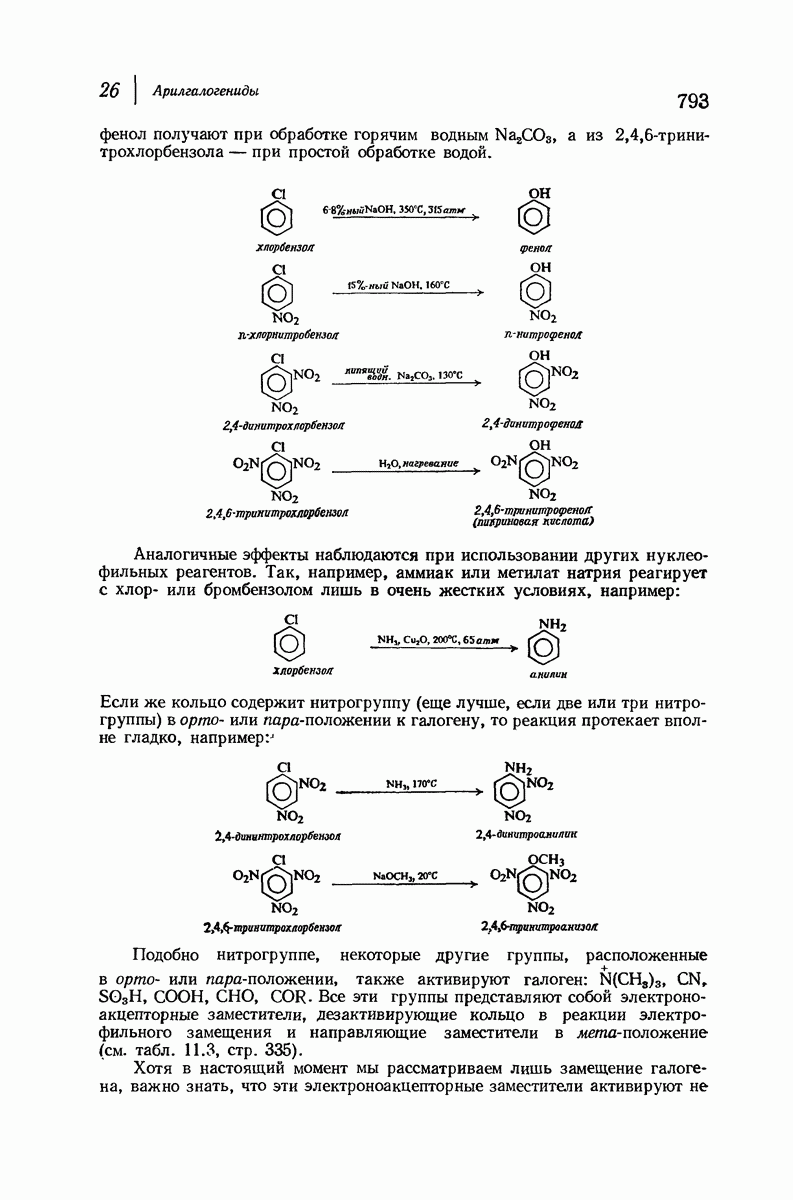
для периодического расстояния, поскольку именно таково расстояние между аминокислотными остатками, измеренное вдоль оси спирали. Для того чтобы уложиться в такую спираль, все аминокислоты должны иметь одну и ту же конфигурацию, что и наблюдается в действительности; кроме того, поскольку они имеют  -конфигурацию, то спираль должна быть правой, как показано на схеме. В настоящее время становится все более очевидным, насколько фундаментальное значение в химии белков имеет структура
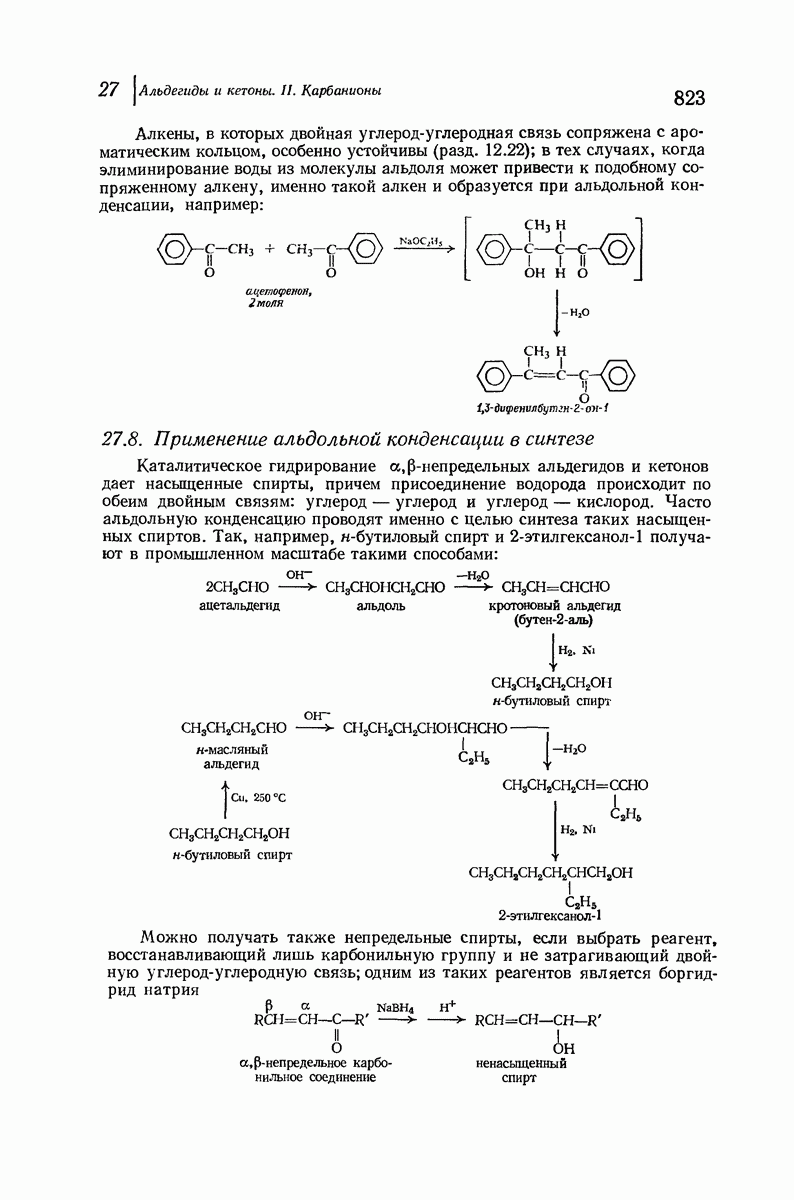
-конфигурацию, то спираль должна быть правой, как показано на схеме. В настоящее время становится все более очевидным, насколько фундаментальное значение в химии белков имеет структура  -спирали.
-спирали.
[Для того чтобы объяснить период идентичности, равный 5,1 А  в
в  -кератине, необходимо рассмотреть то, что является собственно третичной структурой. Полинг предположил, что каждая спираль сама может быть закручена в сверхспираль, имеющую один виток на 35 витков
-кератине, необходимо рассмотреть то, что является собственно третичной структурой. Полинг предположил, что каждая спираль сама может быть закручена в сверхспираль, имеющую один виток на 35 витков  -спирали. Шесть подобных суперспиралей могут быть заплетены вокруг седьмой вытянутой спирали подобно семижильному кабелю.)
-спирали. Шесть подобных суперспиралей могут быть заплетены вокруг седьмой вытянутой спирали подобно семижильному кабелю.)
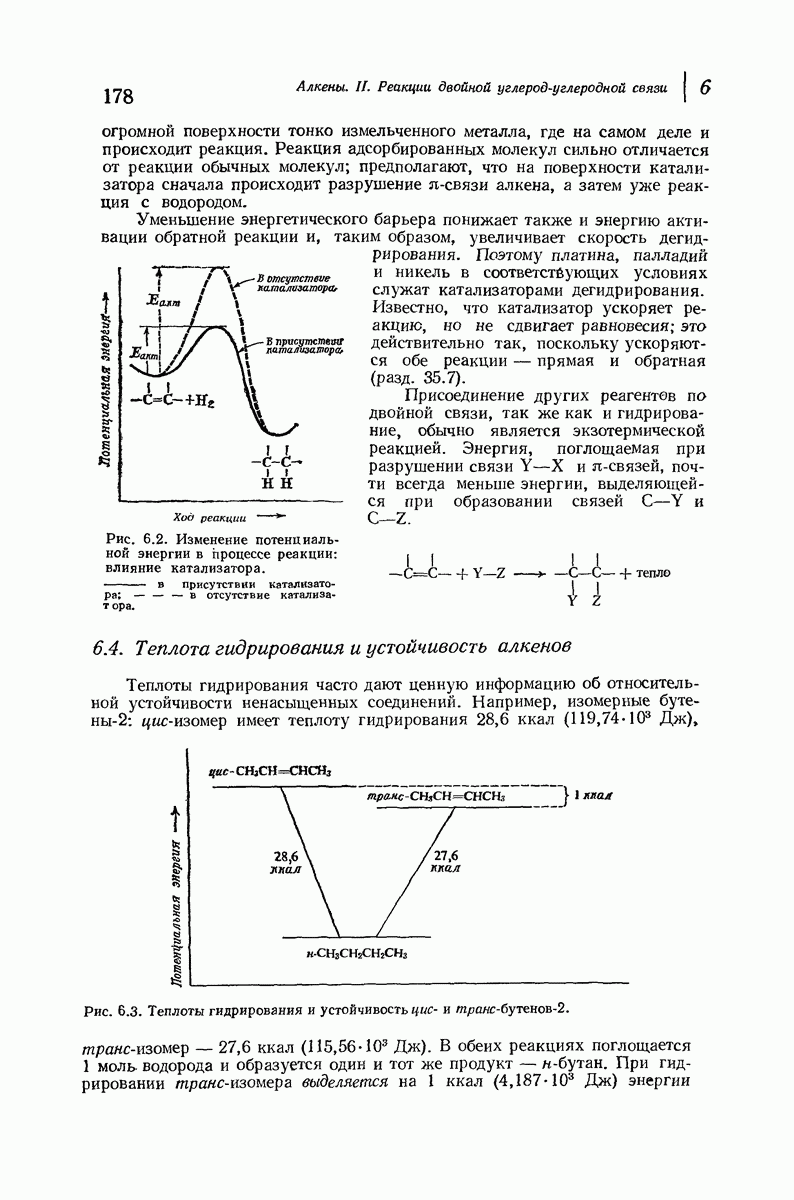
При вытягивании шерсти  -кератин превращается в
-кератин превращается в  -кератин, что сопровождается изменением характера рентгенограмм. Предполагается, что при этом спирали разворачиваются и цепи вытягиваются друг около друга с образованием плоской структуры листа
-кератин, что сопровождается изменением характера рентгенограмм. Предполагается, что при этом спирали разворачиваются и цепи вытягиваются друг около друга с образованием плоской структуры листа  -типа. При этом происходит разрыв водородных связей внутри спирали, и они заменяются водородными связями между соседними цепями. Из-за большего размера боковых групп пептидные цепи менее растянуты период идентичности 6,4 А
-типа. При этом происходит разрыв водородных связей внутри спирали, и они заменяются водородными связями между соседними цепями. Из-за большего размера боковых групп пептидные цепи менее растянуты период идентичности 6,4 А  чем в фиброине шелка [период идентичности 7,0 А
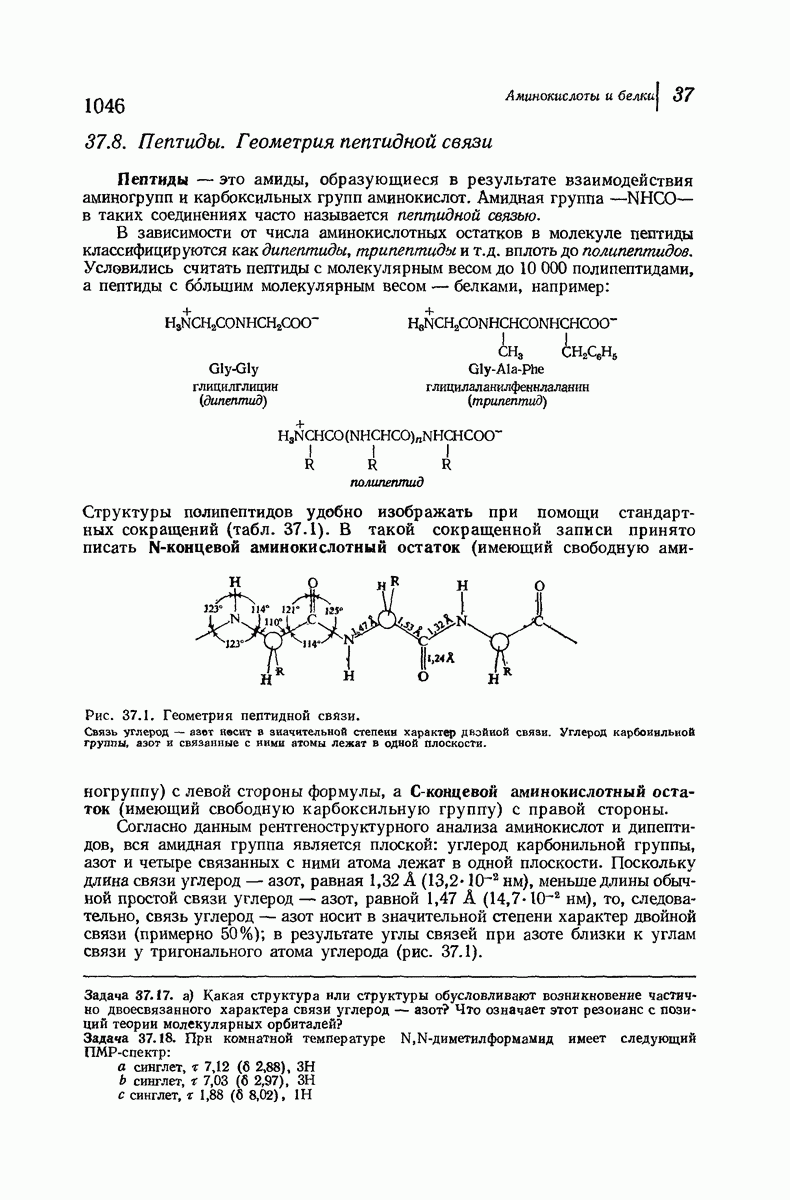
чем в фиброине шелка [период идентичности 7,0 А 
Миозин, фибриллярный белок мышцы, имеет структуру  -спирали. Предполагается, что сокращение мышцы сопряжено с обратимым переходом
-спирали. Предполагается, что сокращение мышцы сопряжено с обратимым переходом  -спирали в
-спирали в  -спираль.
-спираль.
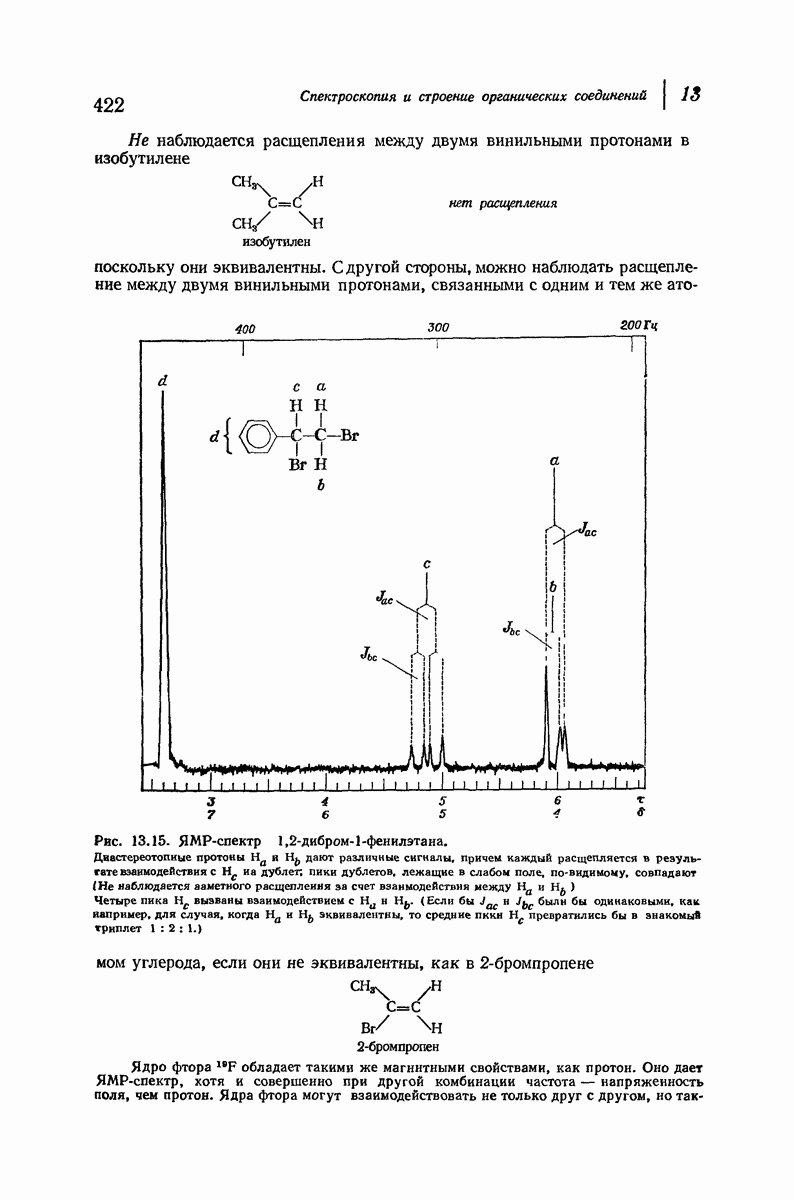
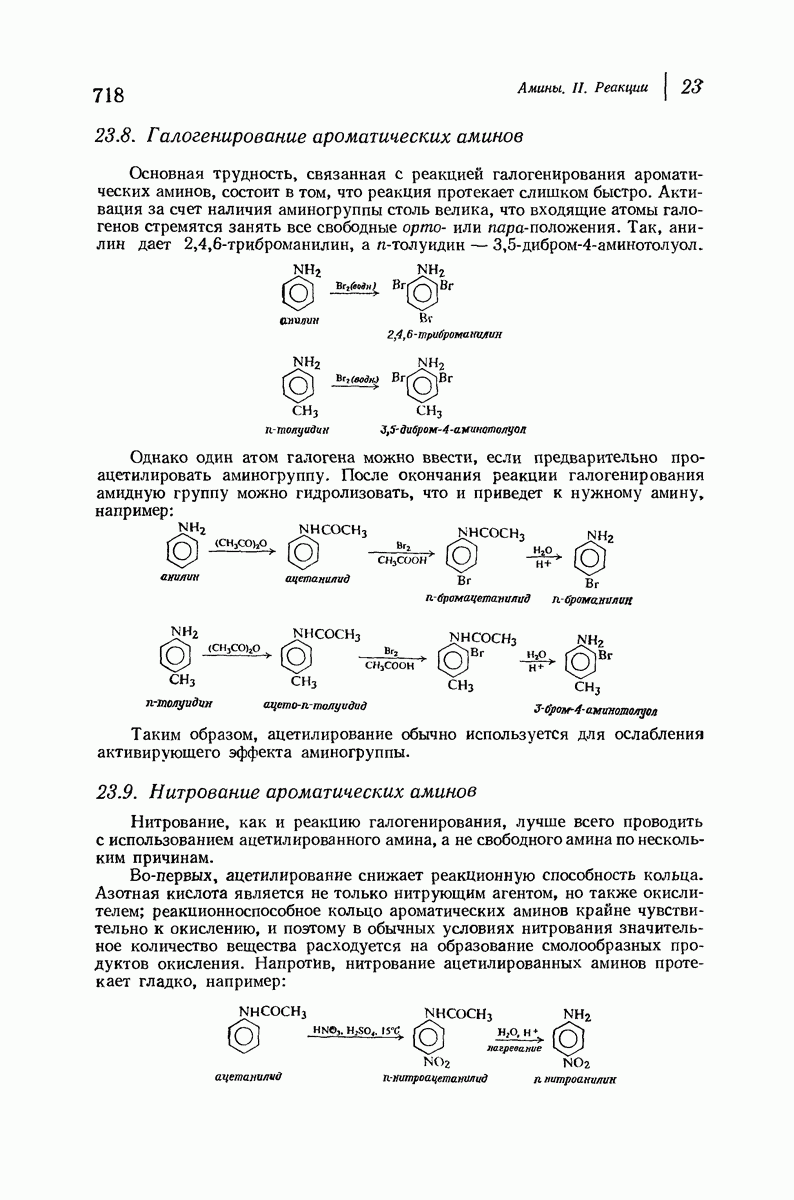
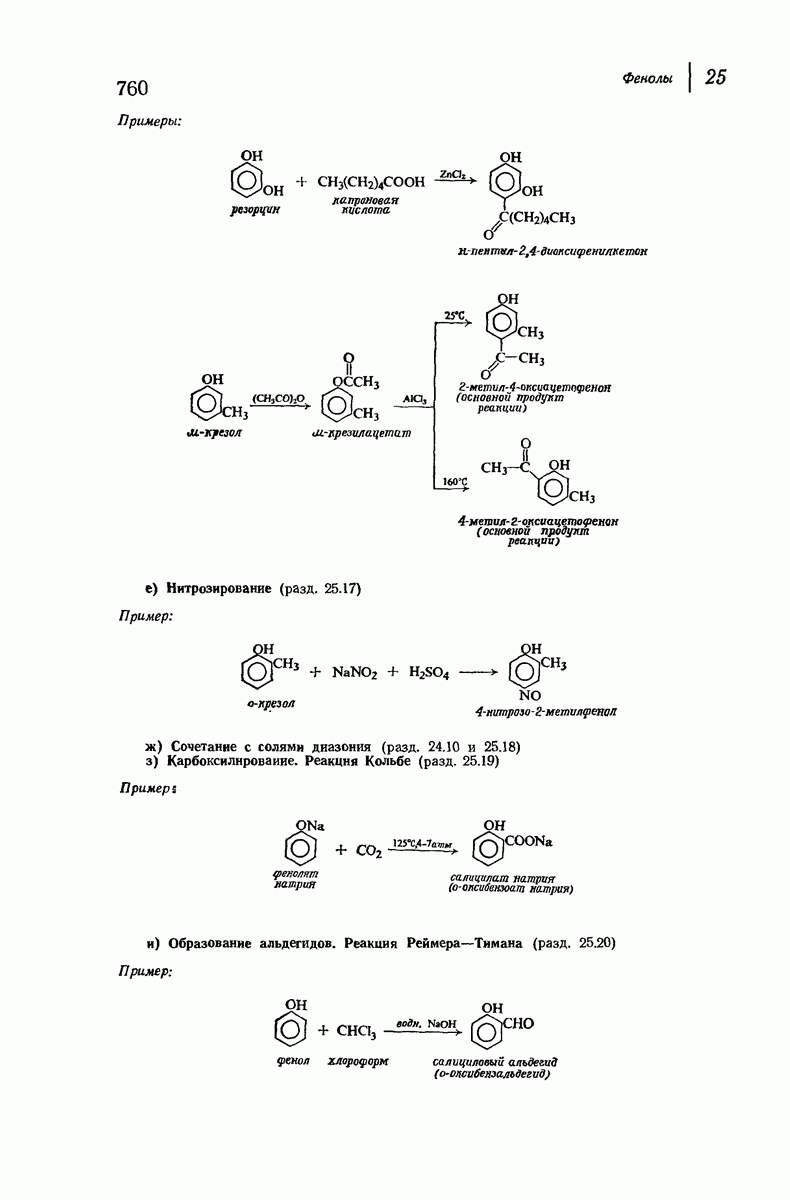
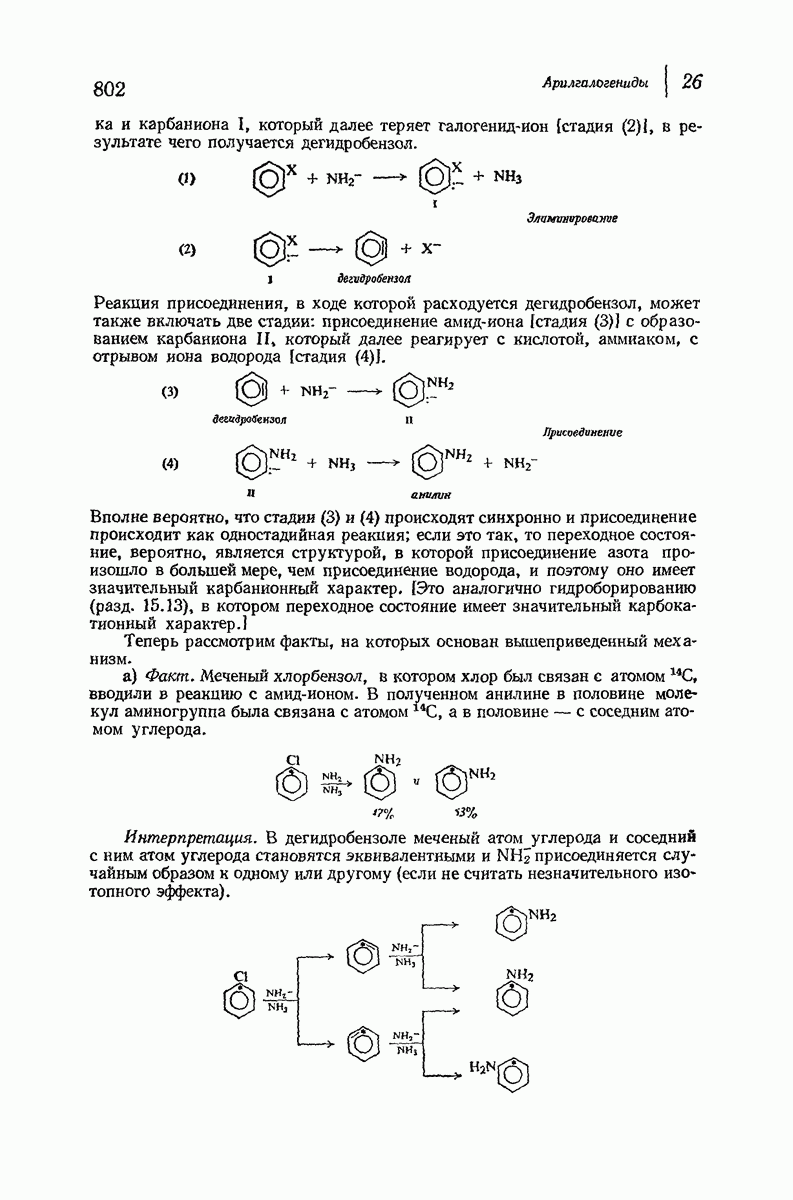
Помимо рентгенограмм, характерных для а- и  -форм белков, известен еще один тип рентгенограмм, а именно для коллагена — белка сухожилий и кожи. На уровне первичной структуры коллаген характеризуется высоким содержанием остатков пролина и оксипролина и частым повторением фрагмента Gly-Pro-Opr. Присутствие пирролидинового кольца в пролине и оксипролине может несколькими способами воздействовать на вторичную структуру. Азот амидной группы не содержит водорода, способного к образованию
-форм белков, известен еще один тип рентгенограмм, а именно для коллагена — белка сухожилий и кожи. На уровне первичной структуры коллаген характеризуется высоким содержанием остатков пролина и оксипролина и частым повторением фрагмента Gly-Pro-Opr. Присутствие пирролидинового кольца в пролине и оксипролине может несколькими способами воздействовать на вторичную структуру. Азот амидной группы не содержит водорода, способного к образованию

водородных связей. Плоское строение пятичленного кольца в сочетании с плоским строением амидной группы не допускает вытягивания пептидной цепи, необходимого для  -структуры, и мешает компактному закручиванию
-структуры, и мешает компактному закручиванию  -спирали.
-спирали.
Структура коллагена сочетает спиральное строение белков  -типа с межмолекулярными водородными связями белков
-типа с межмолекулярными водородными связями белков  -типа. Три пептидные цепи, каждая в форме левой спирали, закручены вокруг друг друга с образованием трехтяжной правой сверхспирали. Небольшие по размеру остатки глицина, находящиеся через каждые два звена в каждой из цепей, образуют пространство, в котором вполне могут разместиться объемистые пирролидиновые кольца двух других цепей. Три цепи удерживаются друг около друга сильными
-типа. Три пептидные цепи, каждая в форме левой спирали, закручены вокруг друг друга с образованием трехтяжной правой сверхспирали. Небольшие по размеру остатки глицина, находящиеся через каждые два звена в каждой из цепей, образуют пространство, в котором вполне могут разместиться объемистые пирролидиновые кольца двух других цепей. Три цепи удерживаются друг около друга сильными
водородными связями между остатками глицина и ОН-группами оксипролина.
При кипячении с водой коллаген превращается в хорошо знакомый растворимый в воде белок желатин; при охлаждении раствор не дает снова коллагена, а образует гель. Молекулярный вес желатина равен одной трети молекулярного веса коллагена. Очевидно, эта обработка приводит к разделению ветвей спирали, разрыву межмолекулярных водородных связей и замене их на водородные связи с молекулами воды.
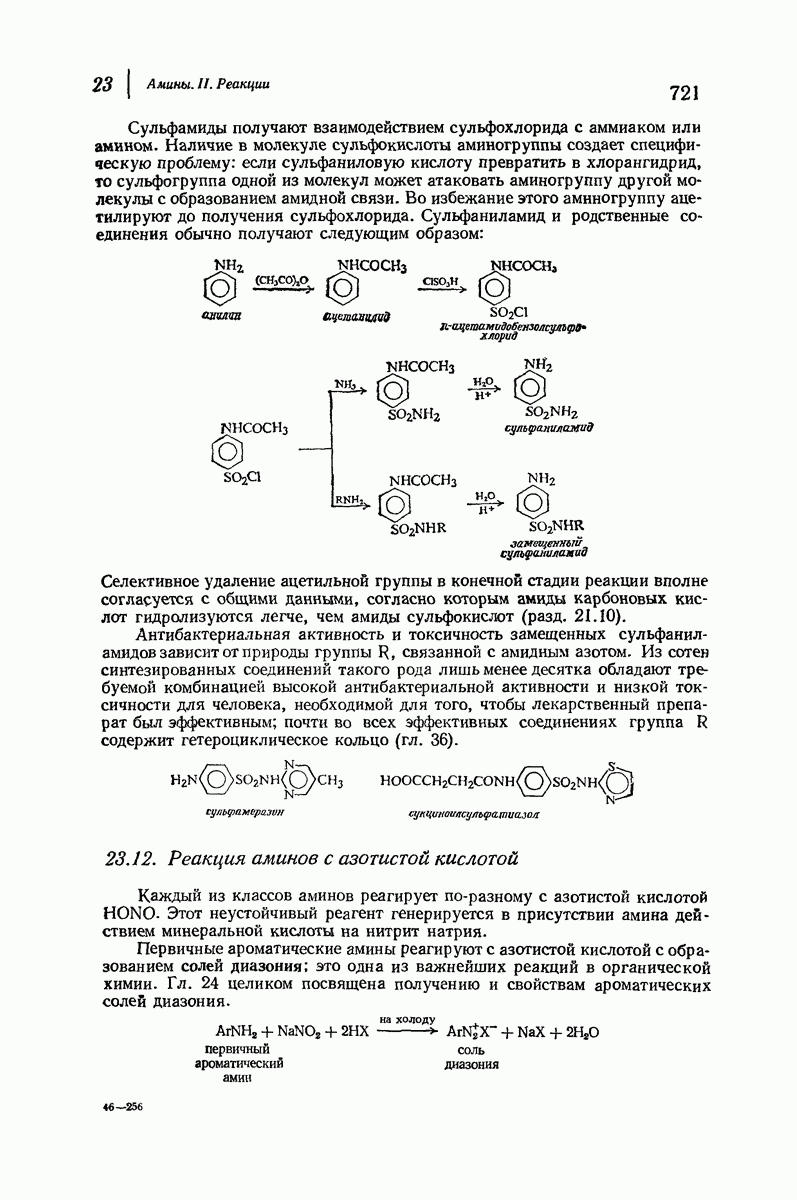
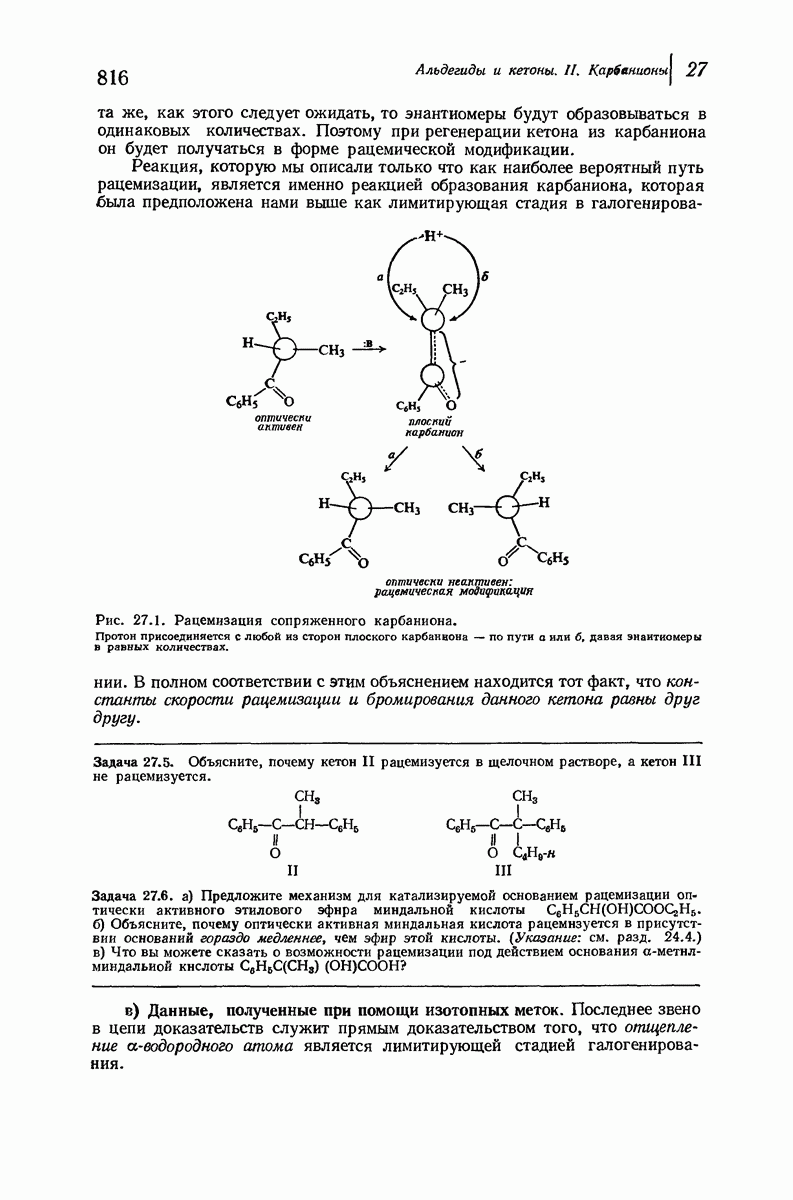
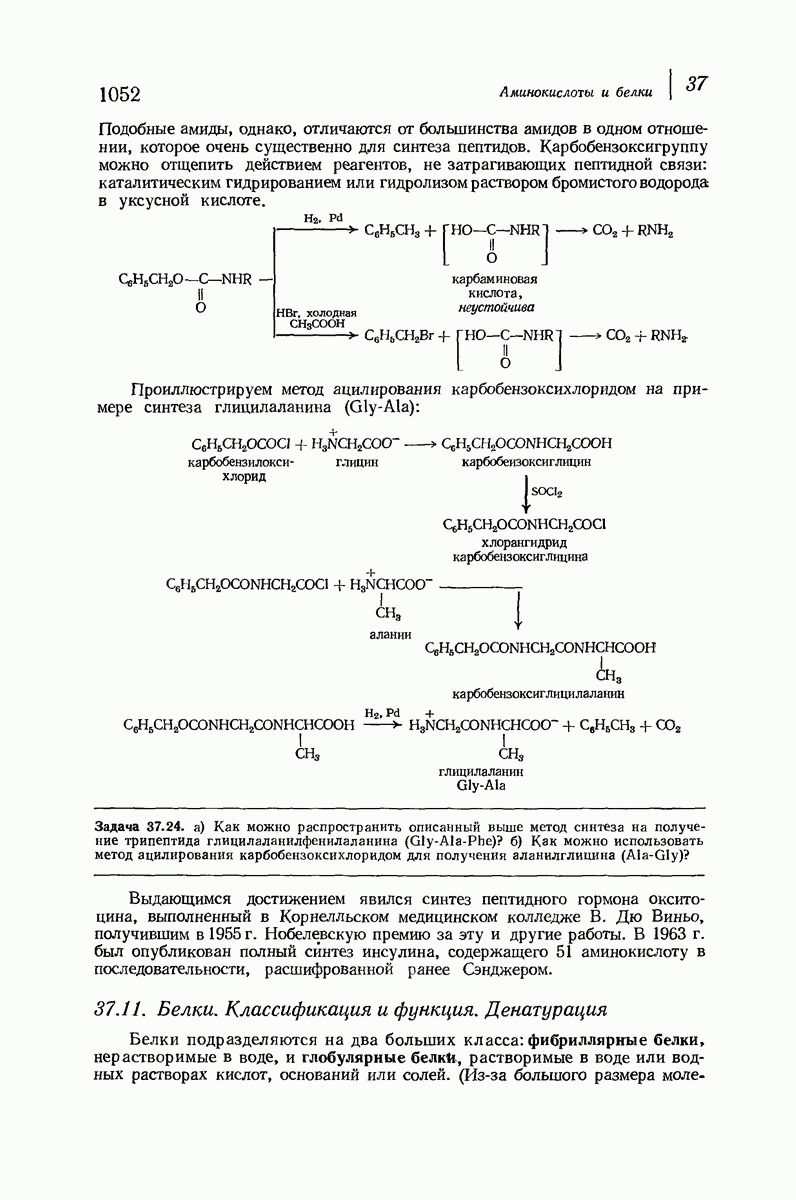
Что касается растворимых глобулярных белков (например, гемоглобина, инсулина, гамма-глобулина, яичного альбумина), то вопрос о характере вторичной структуры еще сложнее. Накапливаются данные, согласно которым и в этом случае  -спираль играет ключевую роль. Подобные длинные пептидные цепи не одинаковы по структуре по всей длине: отдельные их участки свернуты в спирали и являются относительно жесткими; другие участки образуют петли, скручены случайным образом и довольно подвижны. Установлено, что при денатурации белка спиральные участки раскручиваются и цепь в целом приобретает неупорядоченное строение. (Однако опыт показывает, что в определенных условиях раскручивание и возникновение спирали могут быть обратимыми процессами: белок возвращается к исходной вторичной структуре, поскольку это расположение является наиболее стабильным для цепи с данной последовательностью аминокислот.)
-спираль играет ключевую роль. Подобные длинные пептидные цепи не одинаковы по структуре по всей длине: отдельные их участки свернуты в спирали и являются относительно жесткими; другие участки образуют петли, скручены случайным образом и довольно подвижны. Установлено, что при денатурации белка спиральные участки раскручиваются и цепь в целом приобретает неупорядоченное строение. (Однако опыт показывает, что в определенных условиях раскручивание и возникновение спирали могут быть обратимыми процессами: белок возвращается к исходной вторичной структуре, поскольку это расположение является наиболее стабильным для цепи с данной последовательностью аминокислот.)
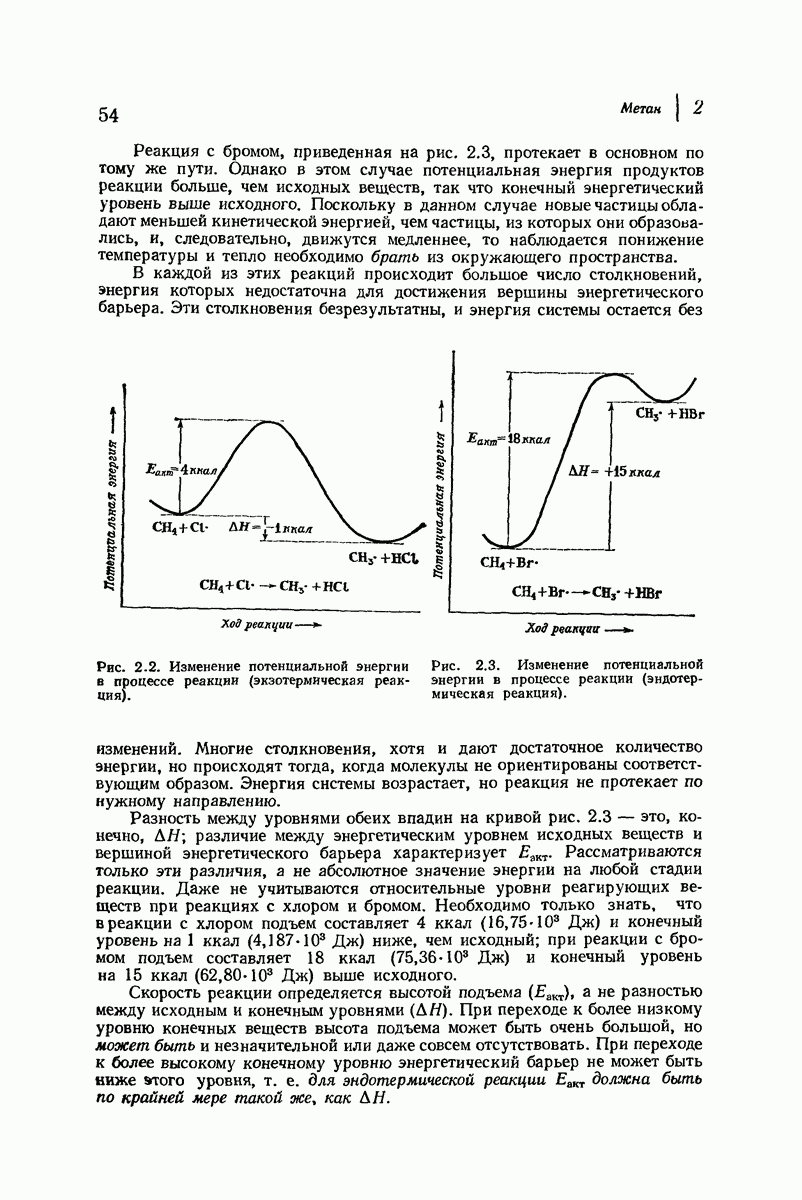
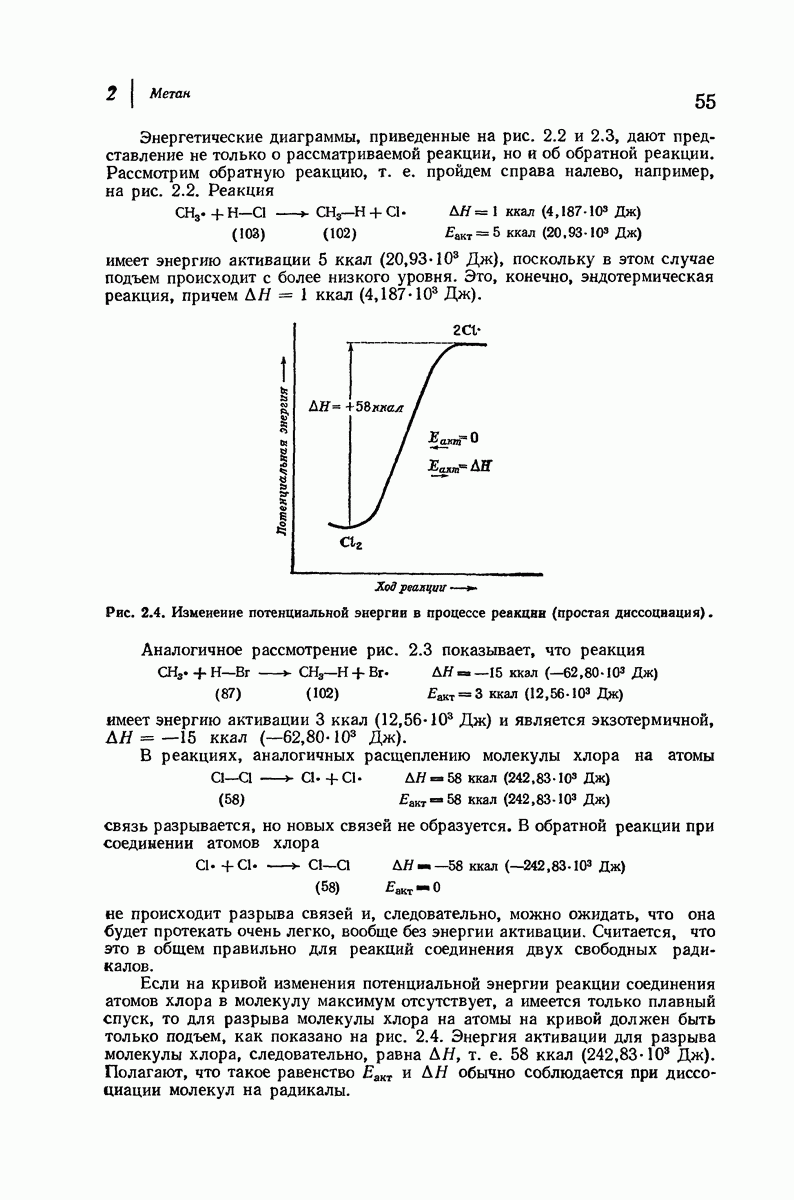
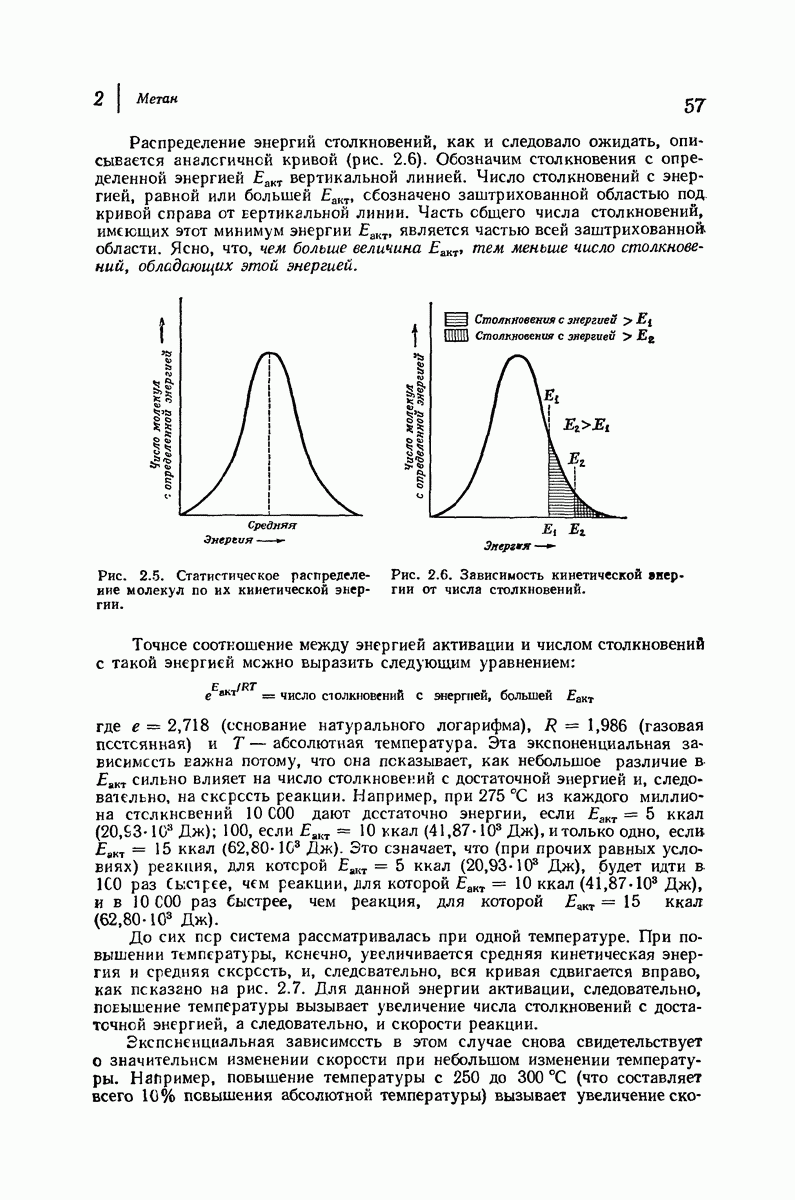
В своих физиологических функциях белки очень высокоспецифичны. Например, фермент может расщеплять  -глюкозиды, но не
-глюкозиды, но не  -глюкозиды или фермент будет отщеплять лишь С-концевые аминокислотные остатки в полипептиде. Биологическая активность белка зависит не только от характера его простетической группы (если она вообще имеется) и данной последовательности аминокислот, а также от формы его молекулы. Как сказал
-глюкозиды или фермент будет отщеплять лишь С-концевые аминокислотные остатки в полипептиде. Биологическая активность белка зависит не только от характера его простетической группы (если она вообще имеется) и данной последовательности аминокислот, а также от формы его молекулы. Как сказал  Фишер в 1894 г.
Фишер в 1894 г.  фермент и глюкозид должны подходить друг к другу, как ключ к замку...».
фермент и глюкозид должны подходить друг к другу, как ключ к замку...».
По-видимому, жесткие спиральные участки в цепи глобулярного белка необходимы для поддержания нужной формы молекулы. Денатурация приводит к раскручиванию спиралей и разрушению характеристичной формы молекулы белка, в результате чего она утрачивает свою специфическую биологическую активность.
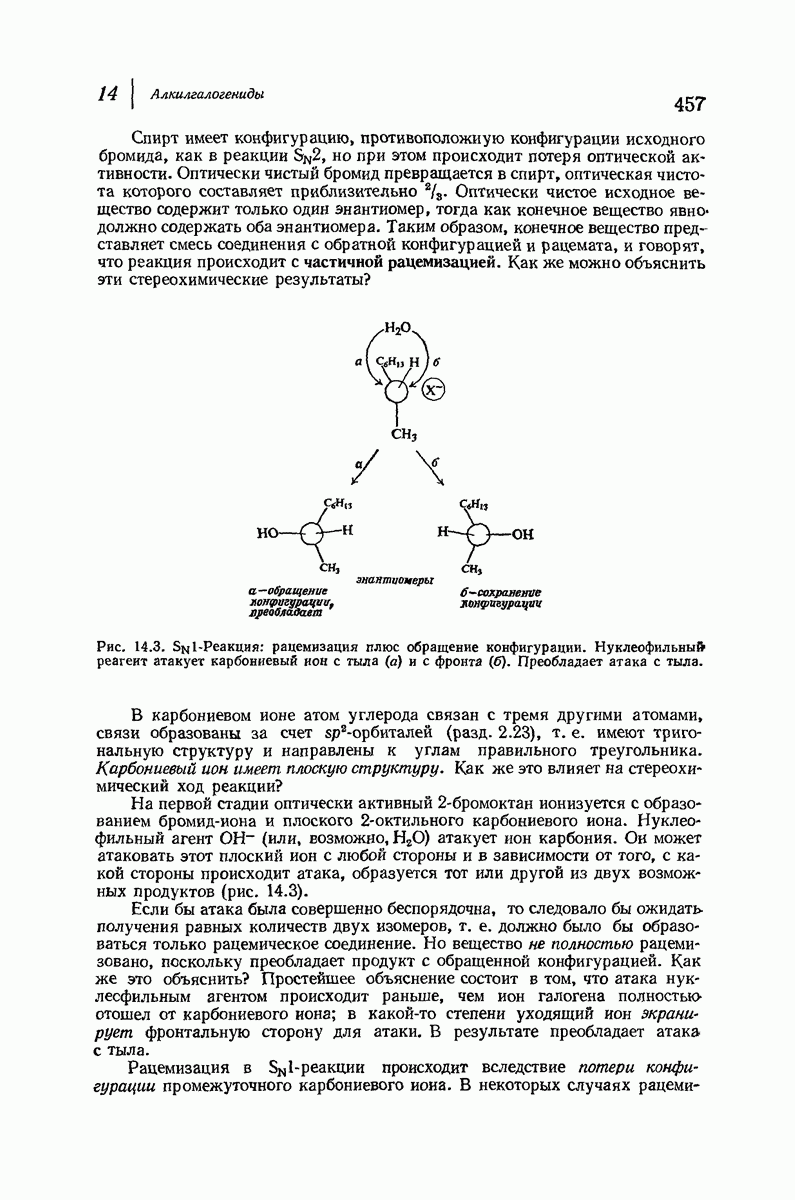
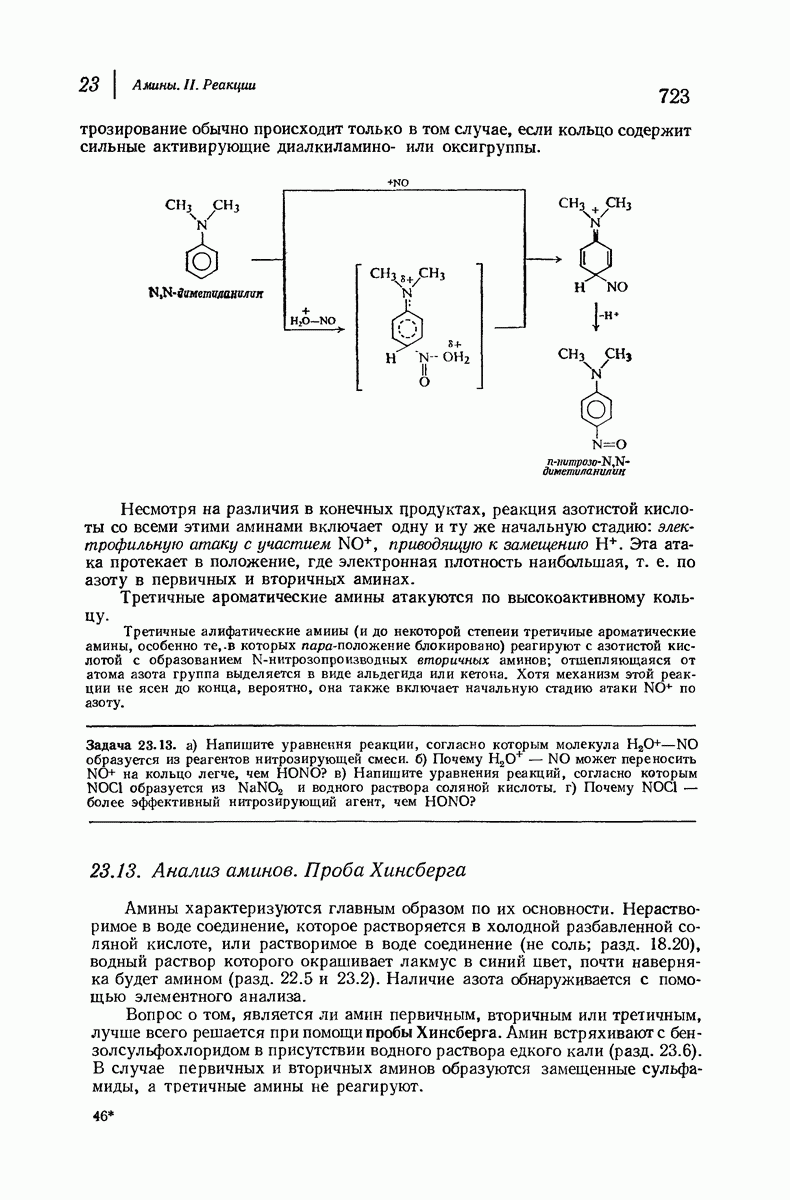
В 1962 г. М. Перутц и Дж. Кендрью (Кембриджский университет) были удостоены Нобелевской премии по химии за работу по установлению структуры гемоглобина и родственного ему миоглобина — молекулы, способной «хранить» кислород. На основании данных рентгеноструктурного анализа и зная аминокислотную последовательность (стр. 1050), они определили трехмерную структуру этих очень сложных молекул: совершенно точно для миоглобина и почти точно для гемоглобина. Они установили, например, что молекула закручена в  -спираль на протяжении шестнадцати звеньев, начиная с концевого N-звена, после чего цепь поворачивает под прямым углом. Исследователи смогли даже сказать, почему она поворачивает: в углу находится звено аспарагиновой кислоты, карбоксильная группа которой нарушает водородные связи, необходимые для продолжения спирали, что и приводит к изменению формы цепи. Четыре сложенные цепи гемоглобина образуют вместе сфероидную молекулу с размерами
-спираль на протяжении шестнадцати звеньев, начиная с концевого N-звена, после чего цепь поворачивает под прямым углом. Исследователи смогли даже сказать, почему она поворачивает: в углу находится звено аспарагиновой кислоты, карбоксильная группа которой нарушает водородные связи, необходимые для продолжения спирали, что и приводит к изменению формы цепи. Четыре сложенные цепи гемоглобина образуют вместе сфероидную молекулу с размерами  Четыре плоские группы гема, каждая из которых содержит атом железа, способный связывать молекулу кислорода, укладываются в отдельных карманах в этой сфере. Когда переносится кислород, то цепи слегка смещаются, в результате чего эти карманы становятся немного меньше по размеру; Перутц описал гемоглобин как «дышащую молекулу». Эти карманы «оторочены» углеводородными остатками аминокислот; подобное неполярное окружение предотвращает перенос электронов между кисдородом и ионом железа и допускает комндексеобразование, необходимое для переноса кислорода.
Четыре плоские группы гема, каждая из которых содержит атом железа, способный связывать молекулу кислорода, укладываются в отдельных карманах в этой сфере. Когда переносится кислород, то цепи слегка смещаются, в результате чего эти карманы становятся немного меньше по размеру; Перутц описал гемоглобин как «дышащую молекулу». Эти карманы «оторочены» углеводородными остатками аминокислот; подобное неполярное окружение предотвращает перенос электронов между кисдородом и ионом железа и допускает комндексеобразование, необходимое для переноса кислорода.